XiaoMi-AI文件搜索系统
World File Search SystemgRNA

低复制CRISPR-CAS13D减轻侧支RNA裂解
g,靶向必需(红色)和非必需(蓝色)基因(n = 4个GRNA)的单个GRNA的归一化耗竭。钻石表示GRNA的中位数。中间95%的非靶向(NT)GRNA的分布以灰色显示。箱图表明所有靶向必不可少的GRNA(平均DEPMAP计时<-1,n = 1,095个细胞系)(红色)和非必需(Chronos> -0.25)(蓝色)基因(蓝色)基因和HAP1细胞中的基因,并使用两侧Mann -Whitney U Test确定了显着性。
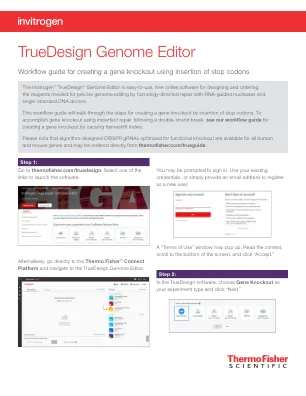
TrueDesign 基因组编辑器
视图将更新以显示“选择 gRNA 选项”。单击适合您需求的 sgRNA 选项 - “预先设计的 sgRNA”(仅限人类和小鼠基因)或“自定义 gRNA 和 TALEN 设计”。如果您对插入终止密码子的区域没有偏好,并且正在人类或小鼠细胞中工作,建议使用“预先设计的 sgRNA”。否则,选择“自定义 gRNA 和 TALEN 设计”。本文档中显示的大多数步骤对于两个工作流程都相似。

纳伊姆,法蒂玛
通过传统育种将新特性引入作物通常需要几十年的时间,但最近开发的基因组序列修饰技术有可能加速这一过程。这些新育种技术之一依赖于 RNA 指导的 DNA 核酸酶 (CRISPR/Cas9) 在体内切割基因组 DNA,以促进序列的删除或插入。这种序列特异性靶向由向导 RNA (gRNA) 决定。然而,选择最佳 gRNA 序列有其挑战。几乎所有当前用于植物的 gRNA 设计工具都是基于动物实验数据,尽管许多工具允许使用植物基因组来识别潜在的脱靶位点。在这里,我们检查了八种不同的在线 gRNA 位点工具的预测一致性和性能。不幸的是,不同算法的排名之间几乎没有共识,排名与体内有效性之间也没有统计学上显着的相关性。这表明,影响植物中 gRNA 性能和/或靶位点可及性的重要因素尚未阐明并纳入 gRNA 位点预测工具中。

CRISPR-Cas9 基因编辑产品 | 简单、快速、精确
与基于质粒的 Cas9 系统相比,完整 RNA 格式的有效载荷更小,可更好地递送到细胞中并提高基因组编辑效率。此外,Cas9 mRNA 可用于具有多个 gRNA 的多重方法。使用此方法可确定哪种 gRNA 序列最适合特定靶标,或通过一次转染编辑多个基因组位点。使用 GeneArt Precision gRNA 合成试剂盒或 GeneArt CRISPR Strings 来制作靶标特异性 gRNA。
![植物生物技术 23.0611a (2023) [adv.pub.]](/simg/8\886d12a05c6e0ea5545c8dfe272b81f8a9782723.webp)
植物生物技术 23.0611a (2023) [adv.pub.]
摘要 马铃薯 (Solanum tuberosum L.) 具有四倍体基因组。要制造缺乏特定基因功能的突变体,必须将突变引入所有四个基因等位基因中。为了实现这一目标,我们开发了一种强大的基因组编辑工具 CRISPR/dMac3-Cas9,它安装了翻译增强子 dMac3,大大增加了下游开放阅读框的翻译。采用三种向导 RNA (gRNA) 的 CRISPR/dMac3-Cas9 系统大大提高了突变的发生率。该系统能够创建颗粒结合淀粉合酶 (GBSS) 和淀粉分支酶 (SBE) 的 4 等位基因突变体。这些突变体显示出功能缺陷的特征,表明我们成功地对马铃薯四倍体基因组进行了有效的基因组编辑。在这里,我们展示了使用 GBSS1 基因突变体时 gRNA 数量对目标基因有效诱变的影响。采用三个 gRNA 基因的 CRISPR/dMac3-Cas9 比采用两个 gRNA 的 CRISPR/dMac3-Cas9 实现了更高的突变效率,表明突变效率受靶区域 gRNA 数量的剂量效应影响。SBE3 基因的等位基因含有导致 gRNA 序列差异的 SNP,但这些 gRNA 有效发挥作用。然而,诱导了许多重排事件和大量缺失。这些结果支持 gRNA 与靶序列准确结合的重要性,这可能为避免脱靶位点的意外突变提供线索。

CRISPR/DCAS9工具包用于玉米基因的功能分析
确实有效。使用基于DCAS9(PDA2),DCAS9-VP64(PDA3)或DCAS9-SRDX(PDA4)载体的CRISPRA/CRISPRI原生质体系统,我们测试了前四个GRNA候选者的有效性,靶向PDS1,CHLH和TRXH和TRXH,FICE(FICE)和3和TRXH,FICE(FICE)和3(FICE)2及3次。与其他关于DCAS9活性的报道类似,我们观察到PDA2在CHLH表达上表现出一些抑制活性(图3A),尽管对于某些GRNA,PDA4的幅度较大(图3B)。用PDA3(CRISPRA构建体)测试,表明TRXH表达增加了两倍,具体取决于GRNA共转染了哪个GRNA(图3C)。这项研究中观察到的GRNA之间的有效性不同,重申了测试多个候选物的必要性。

基于 CRISPR 的配对引导 RNA 抑制 VEGF 治疗脉络膜新生血管
在临床前研究中,利用单个 gRNA 对血管内皮生长因子 A (Vegfa) 进行基于成簇的规律间隔短回文重复序列 (CRISPR) 的基因组破坏可抑制脉络膜新生血管 (CNV),为新生血管性年龄相关性黄斑变性 (AMD) 的长期抗血管生成治疗提供了前景。使用 CRISPR-CRISPR 相关核酸内切酶 (Cas9) 和多个向导 RNA (gRNA) 进行基因组编辑可以通过用基因截断增强插入-缺失 (indel) 突变来增强基因消融效果,但也可能增加脱靶效应的风险。在本研究中,我们比较了腺相关病毒 (AAV) 介导的 CRISPR-Cas9 系统使用单个和配对 gRNA 靶向 Vegfa 基因中在人类、恒河猴和小鼠中保守的两个不同位点的有效性。配对 gRNA 在体外增加了人类细胞中 Vegfa 基因消融率,但在体内并未增强小鼠眼中的 VEGF 抑制。与单个 gRNA 系统相比,使用配对 gRNA 的基因组编辑也显示出相似程度的 CNV 抑制。使用通过测序 (GUIDE-seq) 实现的全基因组无偏双链断裂 (DSB) 识别进行的无偏全基因组分析揭示了由第二个 gRNA 引起的微弱脱靶活性。这些发现表明,使用两个 gRNA 进行体内 CRISPR-Cas9 基因组编辑可能会增加基因消融,但也可能会增加脱靶突变的潜在风险,而针对 Vegfa 基因中的另一个位点作为新生血管性视网膜疾病治疗的功能益处尚不清楚。

使用 GuideScan2 进行全基因组 CRISPR 引导 RNA 设计和特异性分析
基于 CRISPR 的技术已经改变了生命科学,并在治疗学开发中显示出良好的前景 [1],全基因组 CRISPR 筛选通常用于无偏识别各种细胞表型的调节因子。然而,为基于 CRISPR 的基因组扰动设计高效且特异的向导 RNA (gRNA) 带来了计算挑战。不必要的 gRNA 脱靶会导致靶向效率低下以及产生基因毒性,而脱靶信息不完整会导致实验结果的误解 [2]。我们之前开发了 Guide-Scan [3] 用于可扩展的 gRNA 设计,我们和其他人已经证明 GuideScan 在枚举潜在脱靶和估计 gRNA 特异性方面比其他工具更准确 [3, 2]。一个关键的观察结果是,其他 gRNA 设计工具使用的短读比对器虽然对于典型的读取计数量化任务非常有效,但不能详尽地计算次优比对,甚至不能计算多个读取。

Cas-CLOVER™:一种高保真基因组编辑系统...
与使用单个向导 RNA (gRNA) 进行序列特异性引导 CRISPR/Cas9 结合和切割相反,Cas-CLOVER™ 系统使用双 gRNA 引导核酸酶,其中酶的每个半位点亚基都含有催化无活性的 Cas9 (dCas9) 和 IIS 型限制性内切酶 Clo51 的融合蛋白。与广泛用于 TALEN 和锌指核酸酶 (ZFN) 的 FokI 一样,Clo51 活性取决于二聚体的形成,因此 DNA 切割严格依赖于特定距离内两个不同的 gRNA 引导内切酶同时靶向结合。虽然当两个半位点 gRNA 共同递送至细胞时观察到酶的高切割效率,但当单独递送任一半位点 gRNA 时,在原代人类 T 细胞中未观察到靶向破坏。此外,直接比较T细胞中的野生型(WT)CRISPR/Cas9和Cas-CLOVER™表明,在同一基因位点,两种系统都能高效编辑基因组。

grna-seqret:靶向和基因组的通用工具 -
高通量遗传筛选经常用于与表型快速关联并建立序列功能关系。随着CRISPR技术的出现,可以使用合并的指南RNA(GRNA)库和基于测序的测定法对非模型生物进行功能询问以前的遗传性顽固生物,以定量评估并行的每个靶向轨迹。为了帮助构建合并的GRNA组件,我们使用GRNA序列区域提取工具(GRNA-Seqret)开发了用于GRNA选择的硅设计工作流。基于先前开发的CCTOP,GRNA-Seqret启用了针对用户规范区域的GRNA库的自动化,可扩展的设计,或任何原核生物或真核生物的整个基因组。此外,GRNA-Seqret相对于基因或其他特征的任何序列区域的批量提取自动化,有助于插入或缺失构建体的同源臂设计。我们还在计算机中评估了设计的GRNA文库在其他紧密相关的基因组中的应用,并证明对于密切相关的生物体,平均核苷酸同一性(ANI)> 95%> 95%的文库可能是相关的。可以通过https://grna.jgi.doe.gov访问GRNA-Seqret Web应用程序管道。源代码由免费的软件工具和自定义的Python脚本组成,可在https://bitbucket.org/ berkeleylab/grnadesigner/src/master/master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master/Master访问(https://bitbucket.orgelelab/grnadadadeciendiable of。