XiaoMi-AI文件搜索系统
World File Search SystemiPS

使用核转染技术利用 RNP 对人类 iPS 细胞进行 CRISPR 编辑
• 建议戴上手套并使用无核酸酶试管和试剂以避免 RNase 污染。 • 始终保持无菌技术,并使用无菌过滤移液器吸头。 • 所有 EditCo 试剂应根据制造商的建议储存。 • 合成 sgRNA 应溶解在 TE 缓冲液中,并使用无核酸酶水稀释至工作浓度。请参阅 EditCo.com/resources 以查找与溶解和储存合成 sgRNA 相关的最佳实践。 • RNP 可直接在 Nucleofector™ 溶液中形成。 • RNP 复合物在室温下可稳定保存长达 1 小时(可在 4°C 下保存长达一周,或在 -20°C 下保存长达 1 个月)。请注意,在 4°C 下储存的 RNP 可能会在长时间后受到微生物生长的污染。

使用核转染技术利用 RNP 对人类 iPS 细胞进行 CRISPR 编辑
使用 RNA 和 RNP • 建议戴上手套并使用无核酸酶的试管和试剂,以避免 RNase 污染。 • 始终保持无菌技术,并使用无菌过滤移液器吸头。 • 所有 Synthego 和 Nucleofector™ 试剂均应根据制造商的建议储存。 • 合成的 sgRNA 应溶解在 TE 缓冲液中,并使用无核酸酶的水稀释至工作浓度。请参阅 Synthego 快速入门指南,了解溶解和储存合成 sgRNA 的最佳实践。 • RNP 可直接在 Nucleofector™ 溶液中形成。 • RNP 复合物在室温下可稳定保存长达 1 小时(可在 4°C 下保存长达一周,或在 -20°C 下保存长达 1 个月)。请注意,在 4°C 下储存的 RNP 长时间后可能会受到微生物生长的污染。

iPS细胞生成过程中的分子基础及其应用
Shinya Yamanaka 是京都大学 iPS 细胞研究与应用中心 (CiRA) 主任、旧金山格拉德斯通心血管疾病研究所高级研究员和加州大学旧金山分校解剖学教授。Yamanaka 在京都大学 iPS 细胞研究与应用中心 (CiRA) 计划了一项为期五到六年的研究项目,研究诱导多能干细胞 (iPS) 的分子机制和应用。CiRA 聘请了一位年轻的教员 Saito 博士来推动使用基于合成 RNA 的基因操作技术控制细胞命运的研究。他的实验室开发了独特的合成 RNA 分子,以检测和纯化源自 iPS 细胞的靶细胞,并根据细胞内环境控制靶细胞的命运。他负责以下研究项目:开发使用人工 RNA 开关和电路以高安全性和纯度控制哺乳动物细胞命运的新方法。这些 RNA 系统检测靶细胞中表达的特定蛋白质和/或 RNA,然后控制基因表达。
●我们的结果表明IPS是可推广的多...
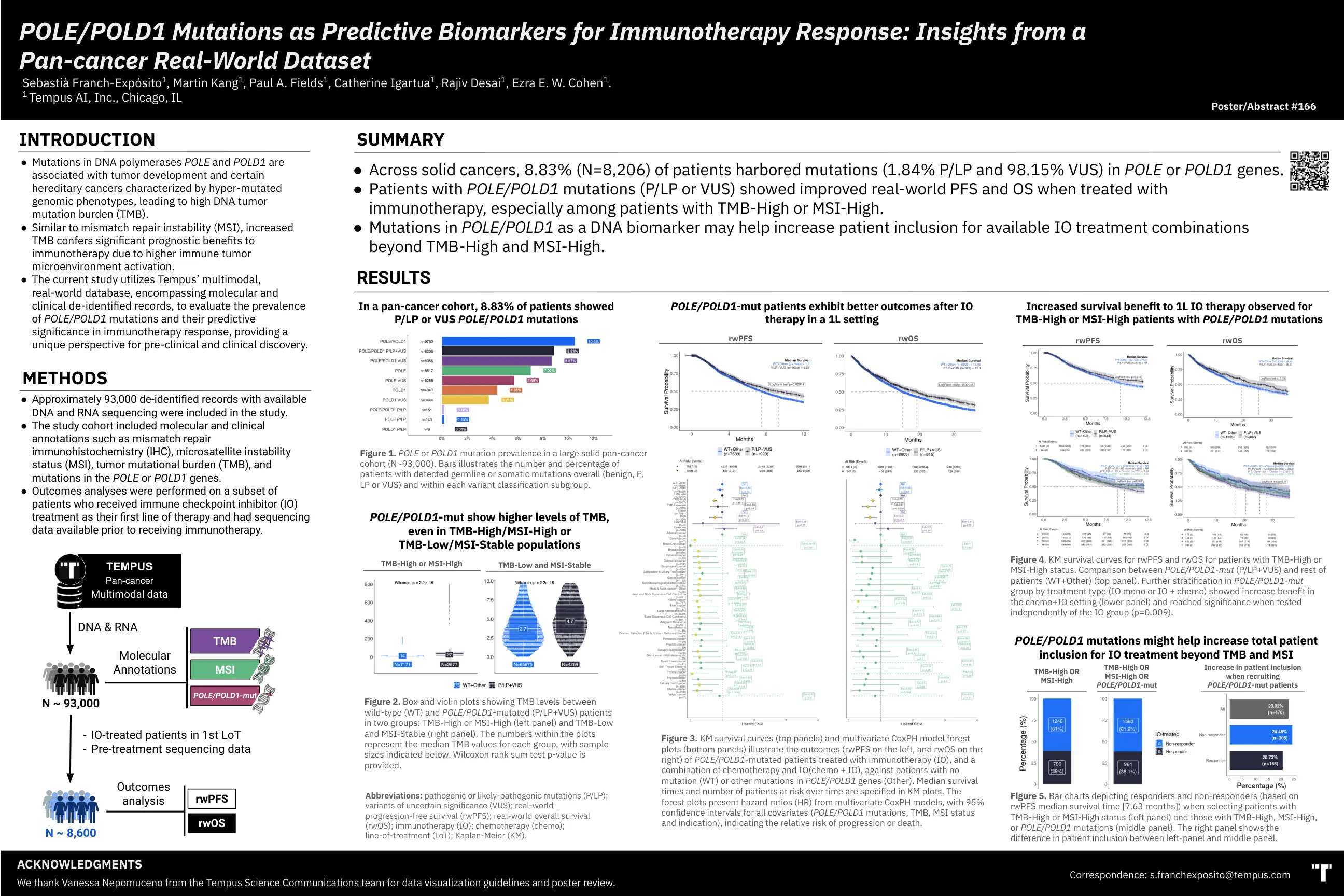
图3。km存活曲线(顶部面板)和多元Coxph森林图(底部面板)说明了POL/POLD 1的左侧的RWPF(左侧RWPF,右侧RWOS)的结果(RWOS),用免疫疗法(IO)治疗的患者(IO)以及与化学疗法和IO + IO + IO + IO + IO + IO + IO(IO)组合的结局(左侧),并与IO + IO + IO(IO)组合进行了突变(基因(其他)。在KM图中指定了随着时间的流逝的中位生存时间和处于危险中的患者人数。森林图具有多元COXPH模型的危险比(HR),所有协变量(POL/POLD1突变,TMB,MSI状态和指示)的置信间隔为95%,表明相对的进展或死亡风险。

利用患者 iPS 细胞衍生的神经干细胞模拟 SHH 驱动的髓母细胞瘤
a 瑞典卡罗琳斯卡医学院微生物学、肿瘤和细胞生物学系 (MTC),邮编 171 65 斯德哥尔摩;b 瑞典乌普萨拉大学生命科学实验室免疫学、遗传学和病理学系,邮编 751 85 乌普萨拉;c 瑞典卡罗琳斯卡医学院神经科学系,邮编 171 65 斯德哥尔摩;d 加拿大多伦多儿童医院发育和干细胞生物学项目,邮编 M5G 0A4;e 加拿大多伦多大学神经外科分部,邮编 M5S 1A8;f 霍普儿童癌症中心 (KiTZ),邮编 69120 海德堡;g 德国癌症研究中心 (DKFZ) 和德国癌症研究联盟 (DKTK) 儿科神经肿瘤学分部,邮编 69120 海德堡; h 分子医学中心分子医学与外科系,卡罗琳斯卡医学院,瑞典斯德哥尔摩 171 76;i 临床遗传学系,卡罗琳斯卡大学医院,瑞典斯德哥尔摩 171 76;j 癌症与衰老研究所,CNRS UMR 7284,INSERM U1081,尼斯索菲亚安提波利斯大学,法国尼斯;k 神经外科分部,临床科学系,斯科讷大学医院,瑞典隆德 221 85;l 神经胶质瘤免疫治疗组,临床科学系,隆德大学,瑞典隆德 221 85;m 妇女与儿童健康系,卡罗琳斯卡医学院,瑞典斯德哥尔摩 171 76

基因组编辑和iPS细胞
图2 利用基因组编辑技术建立疾病模型的研究a:利用源自患有遗传性疾病患者的疾病特异性iPS细胞株,利用基因组编辑技术建立基因修复型iPS细胞株。通过比较两种菌株的受影响细胞类型,我们将分析病理并发现治疗药物。将来,还有望进行通过移植修复型iPS细胞系诱导分化的细胞的基因治疗(细胞治疗)。 b:利用基因组编辑技术将基因突变引入来自健康个体的iPS细胞系,以建立针对疾病的iPS细胞系。通过比较两种菌株的受影响细胞类型,我们将分析病理并发现治疗药物。


使用长ssDNA模板进行同源指导修复的IPS细胞中有效的基因敲击蛋白
CRISPR/CAS技术的常见应用涉及工程基因敲击素,其中DNA序列被取代或插入特定的基因组基因座。In contrast with CRISPR-mediated indels, which result from the error-prone non-homologous end joining (NHEJ) pathway, gene knockins are often engineered via homology-directed repair (HDR), typically through the use of CRISPR reagents (Cas enzyme and guide RNA) in tandem with a DNA template that shares homology with the target site and encodes for the desired modification (Hsu et al., 2014;图1,下面)。用于HDR的模板可以是双链DNA(DSDNA,线性或质粒)或单链DNA(SSDNA),并且最近的发现表明,修复机制取决于使用的模板类型而变化。 dsDNA触发了一种反映减数分裂同源重组(HR)的RAD51依赖性机制,而HDR涉及ssDNA(称为单链模板修复或SSTR)是Rad51独立的,并且需要多个组件,并且需要多个组成部分的Fanconi Anemia Anemia(FA)维修路径(RICHARDARDSON ERATHEWAY(RICHARDARSEN)等。

通过细胞周期和 DNA 修复调节在人类 iPS 细胞中进行协同基因编辑
精准基因编辑旨在生成单核苷酸修饰以纠正或模拟人类疾病。然而,由于效率低下和实用性有限,使用 CRIPSR-Cas9 等核酸酶进行精准编辑的成功率有限。在这里,我们在人类诱导多能干细胞 (iPS) 中建立了荧光 DNA 修复检测,以可视化和量化单等位基因和双等位基因靶向期间 DNA 修复结果的频率。我们发现,通过确定的培养条件和小分子调节 DNA 修复和细胞周期阶段可协同增强同源定向修复 (HDR) 的频率。值得注意的是,在纯合报告细胞中进行靶向可产生高水平的编辑,其中绝大多数为双等位基因 HDR 结果。然后,我们利用高效的双等位基因 HDR 和混合 ssODN 修复模板来产生杂合突变。协同基因编辑是产生人类 iPS 细胞中精确基因修饰的有效策略。

来自人IPS细胞的类器官的功能性肠单层用于药物发现研究
©作者2024。Open Access本文是根据Creative Commons Attribution 4.0 International许可获得许可的,该许可允许以任何媒介或格式使用,共享,适应,分发和复制,只要您对原始作者和来源提供适当的信誉,请提供与创意共享许可证的链接,并指出是否进行了更改。本文中的图像或其他第三方材料包含在文章的创意共享许可中,除非在信用额度中另有说明。如果本文的创意共享许可中未包含材料,并且您的预期用途不受法定法规的允许或超过允许的用途,则您需要直接从版权所有者那里获得许可。要查看此许可证的副本,请访问http://creativecommons.org/licenses/4.0/。Creative Commons公共领域奉献豁免(http://creativecom- mons.org/publicdomain/zero/zero/1.0/)适用于本文中提供的数据,除非在数据信用额度中另有说明。