XiaoMi-AI文件搜索系统
World File Search Systemmediated

在kagome晶格上的电子电子和电子 - phonon介导的配对之间的交叉
我们研究了霍尔斯坦扩展哈伯德模型的电子电子和电子 - phonon介导的配对,并在范霍夫填充物附近的kagome晶格上进行,我们研究了它们对电子配对状态的综合作用。我们发现,它们的组合可以在跨界区域促进异国情调的配对,在该区域中,填充物接近Van Hove的奇异性。尤其是在P型Van Hove填充时,E 1 U(P -Wave)和B 2 U(F Y 3 - 3 - 3 - 3 YX 2 -Wave)配对变得领先,在M -type van Hove flling,E 1 U和A 2 g(I -Wave)配对中得到了促进。此外,我们表明,由于费米表面的sublattice质地,电子 - 光子相互作用获得了显着的动量依赖性,从而可以促进非S波配对。我们对这些配对倾向进行了详细的分析,并讨论了对基于钒的Kagome超导体A V 3 SB 5的含义。


中间层激子介导的BiLayer MOS2中的第二次谐波生成
第二次谐波生成(SHG)是一个非线性光学过程,其中两个光子连贯地组合成两个光子的能量的两倍。的效果SHG。在这里,我们显示了反转对称晶体中非线性光学过程的调整。这种可调节性基于双层MOS 2的独特性能,该特性显示出强烈的光学振荡器强度,但也显示了层间激子的共振。当我们通过改变激光能将SHG信号调谐到这些共振上时,SHG振幅通过几个数量级增强。在谐振情况下,双层SHG信号达到的幅度与单层的两个共振信号相当。在施加的电场中,可以通过鲜明的效应来调节层间激子能量。因此,取消了层间激子退化性,并通过我们的模型计算得出的良好再现了两个数量级,进一步增强了双层SHG响应。
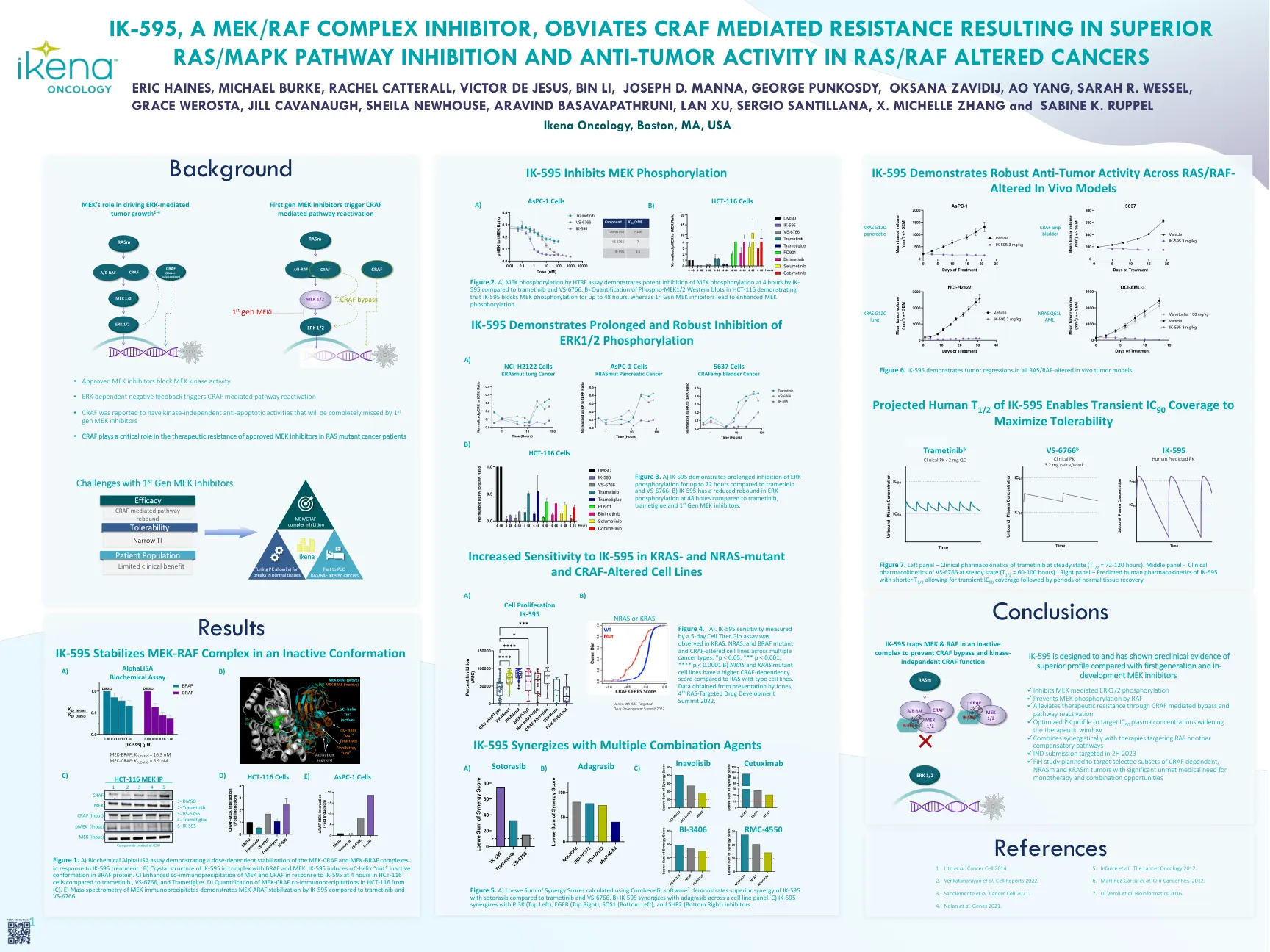
IK-595,一种MEK/RAF复合物抑制剂,避免了CRAF介导的... ik-930,一种用于治疗YAP/TAZ-TEAD依赖性癌症的Tead Paralog选择性抑制剂
图1。a)生化α分析,表明对IK-595处理的MEK-CRAF和MEK-BRAF复合物的剂量依赖性稳定。b)IK-595的晶体结构与BRAF和MEK在复合体中。IK-595在BRAF蛋白中诱导C螺旋“ OUT”无活性构象。 c)与Trametinib,VS-6766和Trametiglue相比,HCT-116细胞在HCT-116细胞中对IK-595的响应增强了MEK和CRAF的共免疫沉淀。 d)(c)中HCT-116中MEK-CRAF共免疫沉淀的定量。 e)MEK免疫沉淀物的质谱法证明了与Trametinib和VS-6766相比,IK-595的MEK-ARAF稳定。IK-595在BRAF蛋白中诱导C螺旋“ OUT”无活性构象。c)与Trametinib,VS-6766和Trametiglue相比,HCT-116细胞在HCT-116细胞中对IK-595的响应增强了MEK和CRAF的共免疫沉淀。d)(c)中HCT-116中MEK-CRAF共免疫沉淀的定量。e)MEK免疫沉淀物的质谱法证明了与Trametinib和VS-6766相比,IK-595的MEK-ARAF稳定。

pt/co中的应变介导的自旋轨道扭矩增强...
Wong,G。D. H.,Xu,Z.,Gan,W.,Ang,C.C.I.,Law,W.C.,Tang,J.,Zhang,W.,Wong,P.K.J.,P.K.J.,Yu,X. 在柔性底物上PT/CO中的应变介导的自旋轨道扭矩增强。 ACS Nano,15(5),8319-8327。 https://dx.doi.org/10.1021/acsnano.0c09404Wong,G。D. H.,Xu,Z.,Gan,W.,Ang,C.C.I.,Law,W.C.,Tang,J.,Zhang,W.,Wong,P.K.J.,P.K.J.,Yu,X.在柔性底物上PT/CO中的应变介导的自旋轨道扭矩增强。ACS Nano,15(5),8319-8327。https://dx.doi.org/10.1021/acsnano.0c09404

转移支原体透明症特异性细胞介导的新生儿猪的免疫力
抽象的支原体溶质膜是猪中enzootic肺炎的主要药物。尽管细胞介导的免疫性(CMI)可能在防御hyopneumoniae的保护中发挥作用,但其从母猪转移到后代的特征很差。因此,在疫苗接种和未接种疫苗的母猪中研究了母体衍生的CMI。还研究了摄入初乳之前的交叉促进对CMI从大坝转移到小猪的转移的潜在影响。六二肠疾病中的hyopneumoniae接种了被人体感染的牛群和47头仔猪,其中24个小猪被交叉寄养,以及三只非疫苗的对照母猪,来自M. hiopneumoniae M. hyopneumoniae-free Herd-free Herd seal-free Herd和24个小猪。疫苗接种的母猪在肌内肌肉内接受了商业细菌,并在for染前6周和3周接受。使用召回测定法评估了不同T细胞子集的TNF-α,IFN-γ和IL-17A的产生。在母猪血液中的细胞因子产生T细胞增加。同样,在这些疫苗接种的母猪中出生的2天大的小猪的血液中检测到了脑性的 - 特异性T细胞。相比之下,在对照母猪的仔猪血液中没有发现脑性的 - 特异性细胞因子产生T细胞。在交叉派生和非交叉式仔猪之间的透明杆菌特异性CMI中没有发现差异。总而言之,不同的Hyopneumoniae M.特异性T细胞子组从母猪转移到后代。需要进一步的研究来研究这些跨性别细胞对小猪中免疫反应的作用及其对透明杆菌感染的潜在保护作用。关键词:支原体溶液,母体免疫,细胞介导的免疫,交叉促进

lbcas12a介导的棉叶卷曲的抑制作用
begomovirus具有传染性,并且严重影响了商业上重要的食物和粮食作物。棉叶卷曲的木木病毒(Clcumuv)是巴基斯坦棉花病毒最主要的特征之一,是对棉花产量的主要限制。目前,植物基因组编辑领域正在通过CRISPR/CAS系统应用(例如基础编辑,主要编辑和基于CRISPR的基因驱动器)进行革命。CRISPR/CAS9系统已成功用于模型和作物植物中的概念概念研究,以针对生物和非生物植物应力。CRISPR/CAS12和CRISPR/CAS13最近已在植物科学中应用于基础和应用研究。在这项研究中,我们使用了一种新型的方法,基于CRRNA的CAS12A工具箱,同时在多个位点靶向Clcumuv基因组的不同ORF。这种方法成功地消除了烟熏本尼亚娜和烟草的症状。从Clcumuv基因组设计了三个单独的CRRNA,针对四个不同ORF(C1,V1和C2和C3重叠区)的特定位点。基于CAS12A的构建体Cas12a-MV是通过金门三向克隆设计的,用于精确编辑Clcumuv Genome。cas12a-MV构建体是通过使用引物UBI-Intron-F1和M13-R1的整个基因组测序来确认的。通过农业纤维化方法,在4周大的尼古蒂亚纳本田植物中进行了瞬态测定。sanger测序表明,CAS12A-MV构建体在病毒基因组的靶位点上产生了相当大的突变。此外,对Sanger测序结果的潮汐分析显示了CRRNA1(21.7%),CRRNA2(24.9%)和CRRNA3(55.6%)的编辑效率。此外,Cas12a-MV构建体通过叶盘方法稳定地转化为烟草Tabacum,以评估转基因植物对Clcumuv的潜力。进行转基因分析,对烟草的转基因植物的DNA进行了PCR,以扩大具有特定底漆的Cas12a基因。传染性克隆在感染性测定中的转基因和非转基因植物(对照)中被农民接种。与具有严重症状的对照植物相比,含有Cas12a-MV的转基因植物表现出少数症状,并且保持健康。与对照植物相比,含有CAS12A-MV的转基因植物显示出病毒积累的显着降低(0.05)(1.0)。结果表明,多重LBCAS12A系统的潜在用途在模型和作物植物中针对贝诺维病毒中发展病毒抗性。

利用纳米载体介导天然药物的癌症治疗策略
目的:核仁素是一种多因素蛋白质,在染色质重塑、mRNA 稳定性、核糖体生物合成、干性、血管生成等方面发挥着重要作用,因此,它是癌症的潜在治疗靶点。本文旨在研究基于多孔硅 (pSi) 纳米载体的天然药物递送系统,以失调的核仁素表达为靶点进行癌症治疗。设计/方法/方法:将槲皮素负载于预先合成和表征的 pSi 纳米粒子中,并研究释放动力学。该研究比较了槲皮素、合成药物阿霉素和负载槲皮素的 pSi 纳米粒子的抑制浓度 (IC50)。此外,用槲皮素处理的乳腺癌细胞系 (MCF-7) 测试了靶基因核仁素的 mRNA 表达。结果:负载槲皮素的 pSi 纳米粒子遵循一级释放动力学。分别在浓度为 312 nM、160 µM 和 50 µM 时测定了针对阿霉素、槲皮素和载有槲皮素的 pSi 纳米粒子的 IC50。结果进一步表明,在用槲皮素处理指数生长的 MCF-7 细胞 48 小时后,核仁素 mRNA 表达下调了 16 倍。研究的局限性/影响:载有槲皮素的 pSi 纳米粒子是否能显著下调核仁素蛋白表达及其对细胞凋亡、细胞增殖和血管生成途径的影响需要进一步研究。实际意义:所提出的基于纳米载体的药物输送系统的实际应用可能为使用天然产物开发针对核仁素失调癌症的靶向治疗铺平了道路,以最大限度地减少传统化疗药物的副作用。原创性/价值:利用天然化合物抑制核仁素和核仁素调节途径并通过纳米载体进行靶向递送尚未完成。关键词:乳腺癌、多孔硅纳米载体、槲皮素;核仁素;靶向治疗本文的参考文献应按以下方式给出:S. Shaw、P. Singh、R. Mishra、R. Singh、R. Nayak、S. Bose,利用纳米载体介导的天然药物的癌症治疗策略,材料与制造工程成就杂志 114/1 (2022) 32-41。DOI:https://doi.org/10.5604/01.3001.0016.1481

NMDAR 介导的 m6A 动态变化与神经元翻译呈负相关
表观转录组修饰在翻译调控中至关重要,对于维持细胞稳态至关重要。N6 甲基腺苷 (m 6 A) 是最丰富且保存良好的表观转录组修饰之一,已知其在神经元功能的各个方面发挥着关键作用。然而,m 6 A 修饰在活动介导的翻译调控和突触可塑性方面的作用尚未研究。在这里,我们研究了 m 6 A 修饰在响应 NMDAR 刺激时的作用。我们一直观察到 5 分钟 NMDAR 刺激会导致 eEF2 磷酸化增加。相应地,NMDAR 刺激在 5 分钟时间点导致 m 6 A 信号显著增加,与整体翻译抑制相关。NMDAR 诱导的 m 6 A 信号增加伴随着 m 6 A 标记 RNA 从翻译到非翻译核糖体池的重新分布。 m 6 A 水平的增加与 NMDAR 刺激下观察到的 FTO 水平降低有很好的相关性。此外,我们还表明,抑制 FTO 可防止 NMDAR 介导的 m 6 A 水平变化。总体而言,我们的结果建立了基于 RNA 的分子读数,它与 NMDAR 依赖的翻译调控相关,有助于理解蛋白质合成的变化。

白垩式介导的呼吸道氧化酶柔韧性控制M.结核病毒力
结果79 80 Chalkophore缺乏结核分枝杆菌上调81对铜剥夺的响应中的呼吸链成分82 83以了解结核分枝杆菌中二甲依替替替替特里利的功能,我们检查了84