XiaoMi-AI文件搜索系统
World File Search Systemmotif

通过机器学习的DNA I -MOTIF的预测-UCL Discovery
摘要i-motifs(IMS)是在富含细胞质的DNA序列中形成的次级str uct uct uct,在基因组中的多个功能中均在v olv中。尽管Putativ e Im forming序列被广泛分布在人类基因组中,但推定的IMS的折叠状态和强度变化了。muc h先前的研究IM已重点是使用生物含量xperiments评估IM折叠特性。ho w e v er,没有专门的计算工具来预测IM结构的折叠状态和强度。在这里,我们介绍了一条机器学习管道,即IM-Weeker,以预测DNA IMS的折叠状态和结构性折叠状态。该程序Im-seeker结合了一个平衡的随机森林分类器,该森林分类器在全基因组IMAB抗体基于基于IMAB的剪切和标记测序数据中训练,以预测折叠状态和极端的梯度增强回归器,以根据文献生物物理数据和我们的内部生物物理实验来估算折叠强度。im-seeker以81%的分类精度预测DNA IM F旧状态,并在测试集上以0.642的确定系数(R 2)估算了F旧强度。模型的解释证实,富含C的序列的核苷酸组成显着影响Im stabilit Y,与含有胞嘧啶和胸腺氨酸的序列具有正相关,并且与鸟嘌呤和腺嘌呤的负相关。

高度重排的非洲假鳃鳉鱼核型中卫星 DNA 基序保守,间质端粒位点缺乏
。CC-BY-NC-ND 4.0 国际许可,根据 提供(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者此版本于 2023 年 3 月 29 日发布。;https://doi.org/10.1101/2023.03.28.534604 doi:bioRxiv 预印本

使用 68Ga-PentixaFor PET 对 690 名实体或血液肿瘤患者的 CXC 基序趋化因子受体 4 表达进行成像
1 德国维尔茨堡大学医院核医学系;2 维也纳医科大学核医学分部,奥地利维也纳;3 日本冈山大学医学、牙科和药学研究生院;4 德国奥格斯堡大学医学院核医学系;5 德国维尔茨堡大学医院内科 II、胃肠病学和 ENETS 卓越中心;6 马里兰州巴尔的摩约翰霍普金斯大学医学院 Russell H Morgan 放射学和放射科学系;7 德国慕尼黑工业大学药物放射化学系;8 德国维尔茨堡 Pentixapharm Würzburg;9 奥地利维也纳医科大学药物化学系;10 德国维尔茨堡大学大学医院医学系 I 内分泌和糖尿病分部; 11 德国维尔茨堡大学医院内科 II、血液学和肿瘤学系。*,# 同等贡献。
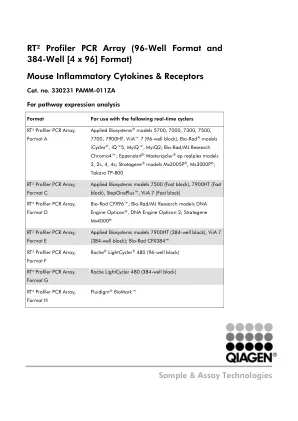
RT² Profiler PCR 阵列(96 孔格式和 384 ... - GeneGlobe
A01 Mm.235137 NM_007926 Aimp1 氨酰 tRNA 合成酶复合物相互作用多功能蛋白 1 A02 Mm.103205 NM_007553 Bmp2 骨形态发生蛋白 2 A03 Mm.1283 NM_011329 Ccl1 趋化因子(CC 基序)配体 1 A04 Mm.4686 NM_011330 Ccl11 趋化因子(CC 基序)配体 11 A05 Mm.867 NM_011331 Ccl12 趋化因子(CC 基序)配体 12 A06 Mm.41988 NM_011332 Ccl17 趋化因子(CC 基序)配体 17 A07 Mm.424740 NM_011888 Ccl19 趋化因子(CC 基序)配体 19 A08 Mm.290320 NM_011333 Ccl2 趋化因子(CC 基序)配体 2 A09 Mm.116739 NM_016960 Ccl20 趋化因子(CC 基序)配体 20 A10 Mm.12895 NM_009137 Ccl22 趋化因子(CC 基序)配体 22 A11 Mm.31505 NM_019577 Ccl24 趋化因子(CC 基序)配体 24 A12 Mm.1282 NM_011337 Ccl3 趋化因子(CC 基序)配体 3 B01 Mm.244263 NM_013652 Ccl4 趋化因子(CC 基序)配体 4 B02 Mm.284248 NM_013653 Ccl5 趋化因子(CC 基序)配体 5 B03 Mm.137 NM_009139 Ccl6 趋化因子(CC 基序)配体 6 B04 Mm.341574 NM_013654 Ccl7 趋化因子(CC 基序)配体 7 B05 Mm.42029 NM_021443 Ccl8 趋化因子(CC 基序)配体 8 B06 Mm.416125 NM_011338 Ccl9 趋化因子(CC 基序)配体 9 B07 Mm.274927 NM_009912 Ccr1 趋化因子(CC 基序) 受体 1 B08 Mm.8021 NM_007721 Ccr10 趋化因子 (CC 基序) 受体 10 B09 Mm.6272 NM_009915 Ccr2 趋化因子 (CC 基序) 受体 2 B10 Mm.57050 NM_009914 Ccr3 趋化因子 (CC 基序) 受体 3 B11 Mm.1337 NM_009916 Ccr4 趋化因子 (CC 基序) 受体 4 B12 Mm.14302 NM_009917 Ccr5 趋化因子 (CC 基序) 受体 5 C01 Mm.8007 NM_009835 Ccr6 趋化因子 (CC 基序) 受体 6 C02 Mm.442098 NM_007720 Ccr8 趋化因子(CC 基序)受体 8 C03 Mm.4861 NM_011616 Cd40lg CD40 配体 C04 Mm.795 NM_007778 Csf1 集落刺激因子 1(巨噬细胞) C05 Mm.4922 NM_009969 Csf2 集落刺激因子 2(粒细胞-巨噬细胞) C06 Mm.1238 NM_009971 Csf3 集落刺激因子 3(粒细胞) C07 Mm.103711 NM_009142 Cx3cl1 趋化因子(C-X3-C 基序)配体 1 C08 Mm.21013 NM_008176 Cxcl1 趋化因子(CXC 基序)配体 1 C09 Mm.877 NM_021274 Cxcl10 趋化因子(CXC 基序)配体 10 C10 Mm.131723 NM_019494 Cxcl11 趋化因子(CXC 基序)配体 11 C11 Mm.303231 NM_021704 Cxcl12 趋化因子(CXC 基序)配体 12 C12 Mm.10116 NM_018866 Cxcl13 趋化因子(CXC 基序)配体 13 D01 Mm.64326 NM_011339 Cxcl15 趋化因子(CXC 基序)配体 15 D02 Mm.4660 NM_009141 Cxcl5 趋化因子(CXC 基序)配体 5 D03 Mm.766 NM_008599 Cxcl9 趋化因子(CXC 基序)配体 9 D04 Mm.234466 NM_009909 Cxcr2 趋化因子(CXC 基序)受体 2 D05 Mm.12876 NM_009910 Cxcr3 趋化因子(CXC 基序)受体 3 D06 Mm.6246 NM_007551 Cxcr5 趋化因子(CXC 基序)受体 5 D07 Mm.3355 NM_010177 Fasl Fas 配体(TNF 超家族,成员 6) D08 Mm.240327 NM_008337 Ifng 干扰素伽马 D09 Mm.379327 NM_008348 Il10ra 白细胞介素10 受体,α

UC Berkeley
单核细胞衍生的巨噬细胞和CD8 + T细胞。The macrophage compartment was heterogeneous and displayed marked enrichment in an inflammatory CCR2 + subpopulation highly expressing Cxcl9 (chemokine [C-X-C motif] ligand 9), Cxcl10 (chemokine [C-X-C motif] ligand 10), Gbp2b (interferon-induced guanylate-binding protein 2b), and Fcgr4 (Fc受体,IgG,低亲和力IV),起源于CCR2 +单核细胞。重要的是,与ICI心肌炎患者相似的表达CXCL9,CXCL10和CD16α(小鼠FCGR4的人类同源物)的巨噬细胞群体(人类的同源物)。暗示了T细胞与CXCL9 + CXCL10 +巨噬细胞之间通过IFN-γ(Interferon Gamma)和CXCR3(CXC趋化因子受体3)信号通路的相互作用。耗尽CD8 + T细胞或巨噬细胞和IFN-γ信号传导的阻断使CXCL9 + CXCL10 +

白质大脑结构可预测语言表现和学习成功
图 2 左半球的网络基序 1 ,其在训练前的流行程度与第 1 天的基线表现相关。 (a)九个大脑区域的矢状面和轴向视图(使用 BrainNet viewer;Xia et al., 2013 创建),其中网络组件具有最高(绝对)连接(标签根据 Glasser et al., 2016 ,在图 b 中)。 (b)网络基序中最强连接的弦图,线条粗细表示基序内连接的绝对权重,颜色表示符号(红色:正,蓝色:负)。请注意,网络基序中具有负权重的连接实际上与基序流行程度呈负相关。然后,绘制 185 个大脑区域中的 67 个,网络基序的主要成分(图 a)以彩色突出显示。 (c)训练前网络流行度(右奇异向量V)与训练第一天受试者在双中心嵌入任务中的表现的回归图(r=.567,p=.046)

screenai:一个用于UI和信息图表的视觉模型
屏幕用户界面(UIS)和信息图表,分享类似的视觉语言和设计原则,在人类通信和人机互动中起重要作用。我们介绍了Screenai,这是一个专门研究UI和信息图表理解的视觉语言模型。我们的模型通过Pix2-Struct的浮雕修补策略改进了Pali体系结构,并通过数据集的独特混合物进行了培训。该混合物的核心是一项新颖的屏幕注释任务,模型必须在其中识别UI元素的类型和位置。我们使用这些文本注释将抄写屏幕屏幕截止到大型语言模型,并通过大规模生成问题索问题(QA),UI导航和摘要培训数据集。我们进行消融研究,以证明这些设计选择的影响。在仅5b个选项中,Screenai在基于UI的和信息图表的任务(乘法DOCVQA,WebSRC和Motif)以及与Simi-lar尺寸的模型相比,在其他基于UI-和信息图表的任务(乘法DOCVQA,WebSRC和Motif)方面取得了新的最先进的结果。最后,我们发布了三个新数据集:一个专注于屏幕注释任务,而两个专注于问题回答的others。

细胞因子 - 武装瘤疱疹单纯疱疹病毒:游戏
摘要细胞因子是调节免疫细胞生长和功能活性的小蛋白质,并且已批准进行癌症治疗。溶瘤病毒是通过直接杀死肿瘤细胞并诱导免疫反应来介导抗肿瘤活性的药物。talimogene laherparepvec是一种单纯性疱疹病毒1型(OHSV),批准用于治疗复发性黑色素瘤,该病毒编码人类细胞因子,粒细胞 - 巨噬细胞 - 巨噬细胞刺激性刺激性刺激因子(GM-CSF)。溶瘤病毒的重要优势是能够将治疗有效载荷运送到肿瘤部位,以帮助促进抗肿瘤免疫。虽然细胞因子作为有效载荷特别有趣,但在溶溶病毒中使用的最佳细胞因子仍然存在争议。In this review, we highlight preliminary data with several cytokines and chemokines, including GM-CSF, interleukin 12, FMS- like tyrosine kinase 3 ligand, tumor necrosis factor α , interleukin 2, interleukin 15, interleukin 18, chemokine (C-C motif) ligand 2, chemokine (C-C motif) ligand 5, chemokine (C-X-C序列4或它们的组合,并显示这些有效载荷如何进一步增强OHSV的抗肿瘤免疫力。OHSV对细胞因子递送的更好了解可以帮助改善癌症患者的溶瘤病毒免疫疗法的临床益处。

构象-Changes-and-atp-Hydrolysy-in-Zika- ...
摘要我们计算研究Zika NS3解旋酶,这是一种使用ATP水解能进行核酸重塑的生物运动。通过经典和QM/MM模拟,我们探索了图案V的构象局势,该构象形象V连接了用于ATP水解和核酸结合的活性位点的保守环。由元磷酸组形成引发的ATP水解涉及由GLU286质子抽象激活的水分子的亲核攻击。基元V氢键通过Gly415骨干NH组与该水键合,从而有助于水解。当无机磷酸盐从镁离子的配位壳移开时,释放自由能,自由能被释放出来,从而诱导了基序V的构象构象构象构象构象形态的显着转移,以在Gly415 NH和Glu285之间建立氢键。Zika NS3解旋酶充当棘轮生物电动机,其基序V转变由Gly415的γ-磷酸在ATPase位点引导。

Libero SoC Linux 环境设置用户指南
6.1. Wind/U 错误:无法连接到服务器上的注册表...............................................................................17 6.2. 警告:未知语言环境...............................................................................................................18 6.3. 警告:无法联系 Web 存储库........................................................................................18 6.4. 错误:无法在 LD_LIBRARY_PATH 中定位 Motif 库.............................................................18 6.5. Wind/U X-toolkit 错误:wuDisplay:无法打开显示.........................................................................19 6.6. Wind/U 错误:发生了致命的注册表 I/O 故障....................................................................................19 6.7. 分段错误“$exedir/$exename”“$@”消息.............................................................................19 6.8. 在 Linux 上运行时,Designer GUI 显得拉伸.............................................................................20 6.9. Libero GUI 在通过 VNC 访问旧版本的 Red Hat 5 时会失真......................................................................20 6.10. 预加载项目时 Libero GUI 无法启动........................................................................................20 6.11. 查看 PDF 文件和在线帮助文件......................................................................................................21 6.12. JRE 库缺失或不兼容......................................................................................................21 6.13. Libero 安装程序在 Red Hat/CentOS 6.x 上停滞.........................................................................................21 6.14. 安装 Linux 软件包时出错....................................................................................................21 6.15. 安装期间出现警告消息:/tmp 磁盘空间不足...............................................................22 6.16. X 库不在 LD_LIBRARY_PATH 中(Libero).............................................................................22 6.17. 缺少 libgthread-2.0.so.0 库(Libero).............................................................................22 6.18.缺少 MOTIF 库 (Libero).....................................................................................................22 6.19. 缺少 libncurses.so.5 库 (ModelSim).....................................................................................22 6.20. 缺少 libXrender 库.......................................................................................................................23 6.21. 缺少 libfontconfig.sol.1 库.............................................................................................................23 6.22. 缺少 libfreetype.so.6 库.............................................................................................................23 6.23. 缺少 libpng.so.6 库.............................................................................................................23