XiaoMi-AI文件搜索系统
World File Search Systempluripotency

间充质干细胞介导肿瘤中的药物递送...
肝细胞癌(HCC)是世界上与癌症相关死亡的第三主要原因。人类羊水间充质干细胞(HAMSC)的表征是多能性,低免疫原性和无肿瘤性的特征。,HAMSC的免疫抑制和抗炎作用使其适合治疗HCC。在这里,我们报告说,通过静脉注射给药的HAMSC通过抑制细胞增殖并用HEPG2细胞诱导肿瘤小鼠的细胞凋亡,从而显着抑制HCC。用GFP标记的HAMSC进行的细胞跟踪实验表明,干细胞具有迁移到肿瘤部位抑制肿瘤生长的能力。重要的是,HAMSC和条件培养基(HAMSC-CM)在体外具有相似的抗肿瘤作用,这表明HAMSC衍生的细胞因子可能参与其抗肿瘤作用。抗体阵列测定法显示,HAMSC高度表达的Dickkopf-3(DKK-3),Dickkopf-1(DKKK-1)和胰岛素样生长因子结合蛋白3(IGFBP-3)。此外,通过抗体的应用或DKK-3,DKK-1和IGFBP-3的特定siRNA的应用进一步证实了HAMSC的抗肿瘤作用。在机械上,HAMSC衍生的DKK-3,DKK-1和IGFBP-3显着抑制了细胞增殖,并通过抑制Wnt/β-catenin信号通路和IGF-1R介导的PI3K/AKT信号通路,促进了HEPG2细胞的凋亡。综上所述,我们的研究表明,HAMSC在体内和体外具有显着的抗肿瘤作用,并且可能在临床上为HCC治疗提供了一种新颖的策略。

患者IPSC中的基因组编辑纠正了最普遍的USH2A突变,并揭示了有趣的突变mRNA表达曲线
遗传性视网膜营养不良(IRD)的特征是进行性光感受器变性和视力丧失。Usher综合征(USH)是一种综合征IRD,其特征是色素性视网膜炎(RP)和听力损失。USH在临床和基因上是异质的,最普遍的病因基因是USH2A。USH2A突变还解释了大量孤立的常染色体隐性RP(ARRP)病例。这种高预期是由于两个经常性的USH2A突变引起的,C.2276G> T和C.2299delg。由于USH2A cDNA的大尺寸,基因增强疗法是无法访问的。但是,CRISPR/CAS9介导的基因组编辑是可行的替代方法。我们使用了增强的链球菌链球菌(ESPCAS9)的特异性CAS9来成功实现诱导多能干细胞(IPSC)患者的两个最普遍的USH2A突变的无缝校正。我们的结果强调了促进ESPCAS9的高目标效率和特种型的功能。一致地,我们没有在校正后的IPSC中识别出任何非靶诱变,这些诱变也保留了多能性和遗传稳定性。此外,对USH2A表达的分析出乎意料地识别了与C.2276G> T和C.229999delg突变相关的异常mRNA水平,这些突变在校正后恢复。综上所述,我们有效的CRISPR/CAS9介导的USH2A突变校正策略为USH和ARRP患者提供了潜在治疗的希望。

导航干细胞培养:见解,技术,挑战和前景
干细胞研究对再生医学和疾病建模具有巨大的希望,使对干细胞培养的理解和优化成为推进这些治疗应用的关键方面。这项全面的综述提供了对干细胞培养的深入概述,包括一般信息,当代技术,遇到的问题和未来的观点。本文首先解释了各种干细胞类型的基本特征,从而阐明了适当的培养条件在维持多能或谱系承诺中的重要性。对既定文化技术的详细探索阐明了干细胞培养方法不断发展的景观。常见的挑战,例如遗传稳定性,异质性和差异性效率,并深入讨论了旨在解决这些障碍的尖端策略和技术的见解。此外,本文深入研究了基材材料,培养基成分和生物物理线索对干细胞行为的影响,强调了微环境和细胞命运决策之间的复杂相互作用。随着干细胞研究的发展,道德考虑和监管框架变得越来越重要,从而在文化实践的背景下促使对这些方面进行了批判性检查。最后,本文探讨了新兴的观点,包括将人工智能和机器学习在优化文化条件中的整合,以及干细胞衍生产品在个性化医学中的潜在应用。这个全面的概述旨在为研究人员和临床医生提供宝贵的资源,从而更深入地了解干细胞培养物及其在推进再生医学和生物医学研究中的关键作用。

从非合成性USH2A相关性视网膜炎的患者
众所周知,这是由于USH2A中存在“视网膜特异性”等位基因的原因。因此,出现至少一份视网膜特异性等位基因副本的患者将出现孤立的RP表型。高度复发的c.2276g> t突变是视网膜特异性等位基因(Lenassi等,2015)。在这里,我们报告了IPSC系列的生成,该患者从ARRP出现的患者和复合het erozygous c.2276g> t等位基因和另一个错位等位基因,C.7352 t> c,据我们所知,以前是未报告的。从患者皮肤活检中分离出人真皮成纤维细胞,并使用非整合性细胞调整-IPS 2.0 sendai重编程套件对重编程进行了重新编程。该套件包含仙台病毒(SEV)载体汽车,将OCT3/4,SOX2,KLF4和C-MYC转基因赋予了多脂能力。转变后三到四个星期,基于形态标准单独选择了类似IPSC的菌落。选择了Inmi005-A IPSC系列以进一步特征,因为它具有典型的IPSC菌落形态,该形态的紧密堆积的小细胞被尖锐的边界包围(图1 a)。使用逆转录(RT)-PCR确定外源载体的损失(图1 b)。通道12(p12)在SEV基因组和KLF4,KOS和C-MYC Cassettes的RT-PCR结果为阴性,类似于非转导的成纤维细胞,用作阴性对照。相比之下,被用作阳性对照的转导的成纤维细胞(纤维 + SEV)对四个靶标呈阳性。我们进行了

开发用于高内涵筛选的 CRISPR/Cas9 介导荧光报告基因人类多能干细胞系
摘要:应用 CRISPR/Cas9 系统将荧光蛋白敲入人类多能干细胞 (hPSC) 中的内源性目的基因,有可能促进基于 hPSC 的疾病建模、药物筛选和移植疗法优化。为了评估荧光报告 hPSC 系用于高内涵筛选方法的能力,我们将 EGFP 靶向内源性 OCT4 基因座。产生的 hPSC–OCT4–EGFP 系表达与多能性标记物一致的 EGFP,并且可以适应多孔格式以进行高内涵筛选 (HCS) 活动。然而,在长期培养后,hPSC 暂时失去了 EGFP 表达。或者,通过将 EGFP 敲入 AAVS1 基因座,我们建立了稳定且一致的 EGFP 表达 hPSC–AAVS1–EGFP 系,该系在体外造血和神经分化期间保持 EGFP 表达。因此,hPSC–AAVS1–EGFP 衍生的感觉神经元可适应高内涵筛选平台,该平台可应用于高通量小分子筛选和药物发现活动。我们的观察结果与最近的发现一致,表明在 OCT4 基因座进行 CRISPR/Cas9 基因组编辑后会出现高频率的靶向复杂性。相反,我们证明 AAVS1 基因座是 hPSC 中的安全基因组位置,具有高基因表达,不会影响 hPSC 质量和分化。我们的研究结果表明,应应用 CRISPR/Cas9 整合的 AAVS1 系统来生成稳定的报告 hPSC 系以用于长期 HCS 方法,并且它们强调了仔细评估和选择应用的报告细胞系以用于 HCS 目的的重要性。
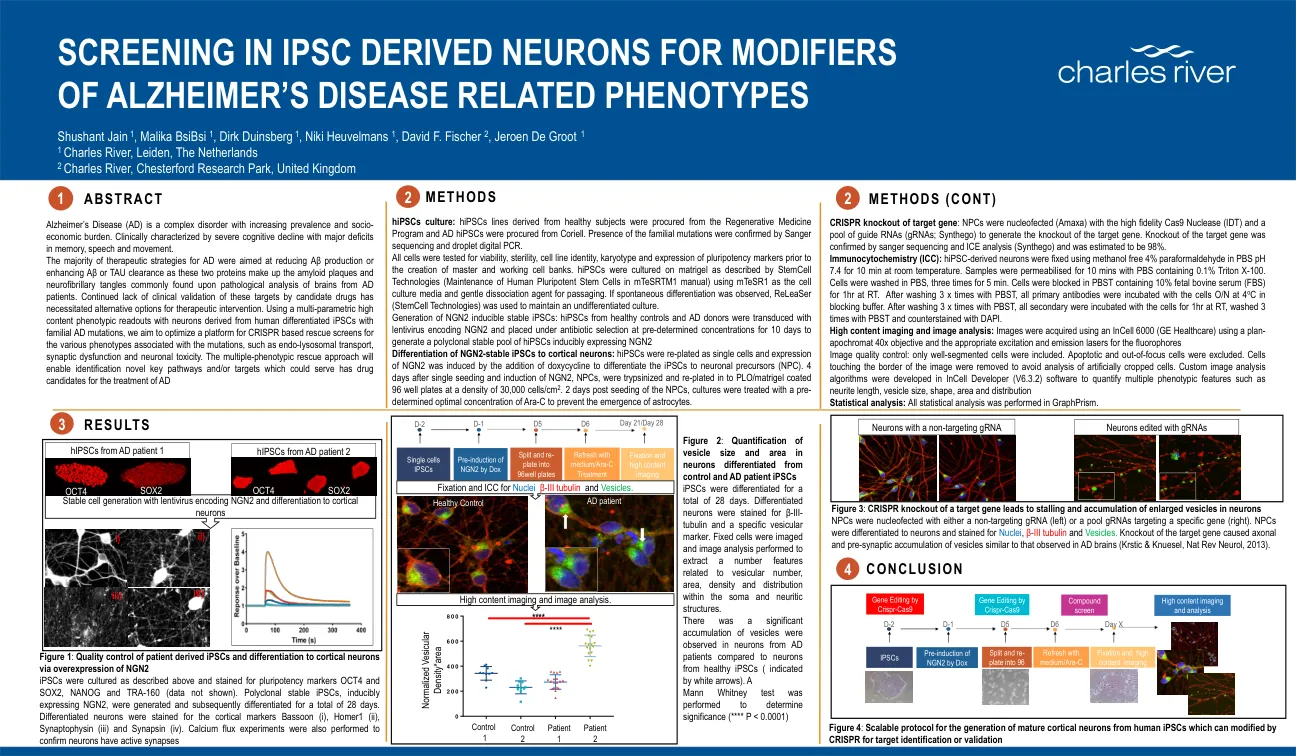
在 IPSC 衍生的神经元中筛选阿尔茨海默病相关表型的修饰因子 | Charles River
hiPSC 培养:来自健康受试者的 hiPSC 系来自再生医学计划,AD hiPSC 来自 Coriell。通过 Sanger 测序和液滴数字 PCR 确认存在家族性突变。在创建主细胞库和工作细胞库之前,对所有细胞进行了活力、无菌性、细胞系身份、核型和多能性标志物表达测试。hiPSC 在基质胶上培养,如 StemCell Technologies(mTeSRTM1 手册中的人类多能干细胞维护)所述,使用 mTeSR1 作为细胞培养基和温和的解离剂进行传代。如果观察到自发分化,则使用 ReLeaSer(StemCell Technologies)来维持未分化培养。 NGN2 诱导的稳定 iPSC 的生成:健康对照和 AD 供体的 hiPSC 被编码 NGN2 的慢病毒转导,并在预定浓度的抗生素选择下放置 10 天,以生成可诱导表达 NGN2 的 hiPSC 多克隆稳定池。NGN2 稳定的 iPSC 向皮质神经元的分化:将 hiPSC 重新接种为单细胞,并通过添加强力霉素诱导 NGN2 的表达,以将 iPSC 分化为神经元前体 (NPC)。单次接种和诱导 NGN2 4 天后,将 NPC 用胰蛋白酶消化并重新接种到 PLO/matrigel 包被的 96 孔板中,密度为 30,000 个细胞/cm 2 。在 NPC 接种 2 天后,用预定的最佳浓度的 Ara-C 处理培养物以防止星形胶质细胞的出现。

胚胎DNA甲基化程序通过CTCF拮抗作用调节顺式调节景观
1。巴黎大学,CNRS,Institut Jacques Monod,F-75013法国2。CEA,CEA,CNRS,CNRS综合生物学研究所(I2BC),法国F-91998 GIF-SUR-YVETTE,法国 *这些作者同样贡献了†对应:Maxim.greenberg@ijm.fr摘要,在哺乳动物的胚胎生成期间,两种甲基元素(5-Cyylimens)(5-Cyylimens)( (3D)在称为“表观遗传重编程”的过程中对染色质结构进行了深刻的重塑。表观遗传重编程的一个研究的方面是5mec通量本身如何影响3D基因组。 鉴于DNA结合对染色体折叠的关键调节剂的5mec-敏感性:CTCF。 我们使用小鼠胚胎干细胞(ESC)分化方案对CTCF结合景观进行了介绍,该方案模拟了幼稚多能的退出,其中全局DNA甲基化水平在四天内开始较低,并在四天内提高到躯体水平。 我们利用了这一事实,即缺乏DNA甲基化机制的小鼠ESC表现出全球相似的分化动力学,从而使CTCF不正调对基因表达的更微妙作用进行解剖。 我们通过在野生型和突变条件下进行CTCF HICHIP来评估异常的CTCF-CTCF接触,在没有5mec的情况下。 ,鉴于H3K27AC在主动启动子和增强子上富含H3K27AC,我们继续评估了错误调节的CTCF结合对顺式调节接触的影响。 使用DNA甲基化表观组编辑,我们能够直接证明DNA甲基马克能够影响CTCF结合。CEA,CEA,CNRS,CNRS综合生物学研究所(I2BC),法国F-91998 GIF-SUR-YVETTE,法国 *这些作者同样贡献了†对应:Maxim.greenberg@ijm.fr摘要,在哺乳动物的胚胎生成期间,两种甲基元素(5-Cyylimens)(5-Cyylimens)( (3D)在称为“表观遗传重编程”的过程中对染色质结构进行了深刻的重塑。表观遗传重编程的一个研究的方面是5mec通量本身如何影响3D基因组。鉴于DNA结合对染色体折叠的关键调节剂的5mec-敏感性:CTCF。我们使用小鼠胚胎干细胞(ESC)分化方案对CTCF结合景观进行了介绍,该方案模拟了幼稚多能的退出,其中全局DNA甲基化水平在四天内开始较低,并在四天内提高到躯体水平。我们利用了这一事实,即缺乏DNA甲基化机制的小鼠ESC表现出全球相似的分化动力学,从而使CTCF不正调对基因表达的更微妙作用进行解剖。我们通过在野生型和突变条件下进行CTCF HICHIP来评估异常的CTCF-CTCF接触,在没有5mec的情况下。,鉴于H3K27AC在主动启动子和增强子上富含H3K27AC,我们继续评估了错误调节的CTCF结合对顺式调节接触的影响。使用DNA甲基化表观组编辑,我们能够直接证明DNA甲基马克能够影响CTCF结合。最后,对印迹ZDBF2基因的详细解剖表明,CTCF的5mec-抗抗酸如何允许分化过程中适当的基因调节。这项工作提供了全面的概述,概述了DNA甲基化如何影响早期胚胎事件的相关模型中的3D基因组。

干细胞研究
转基因株系采用第二代 CRISPRa 系统,该系统携带与异源三聚体 VPR 反式激活因子融合的核酸酶缺陷型 dCas9,该异源三聚体 VPR 反式激活因子由 VP64、p65 和 RTA 结构域组成。该系统可用于解释任何所需细胞类型的内源性调控机制。使用基于 CRISPR/Cas9 的基因组编辑方法,我们以 AAVS1 人类基因组位点为目标,分别引入先前描述的 dCas9VPR-tdTomato(Schoger 等人,2020 年)和嘌呤霉素盒,这些盒受 CAG 和 EF1a 启动子的控制(图 1 A)。采用优化的核转染方案转染 LhiPSC-GR1.1 细胞。转染后,选择具有 tdTomato 表达的细胞并通过 PCR 进行基因分型(图 1B,引物结合如图 1A 所示,黑色引物仅扩增野生型 (WT) 片段;绿色引物扩增插入的构建体)。随后,扩增、分析和冷冻保存两个阳性克隆(#2 和 #3)。DNA 测序数据证实了 AAVS1 基因座中的正确和纯合敲入转基因整合(图 1C,显示为克隆#2)。PCR 结果显示,在筛选的 15 个克隆中,11 个克隆含有纯合插入(命名为 CRISPRa 细胞),1 个克隆是杂合的,3 个克隆不含有插入而是含有 WT 完整基因座(用作对照细胞)(数据未显示)。通过分析 PCR 和测序预测的前五个脱靶位点进行脱靶分析;在这些位点中均未发现任何编辑事件。对照电穿孔和非电穿孔 (参考) 系用于比较 (补充图 1A)。所有系的支原体检测均为阴性。通过基于 SNP 的核型分析和标准 G 带证明了 CRISPRa 克隆 #2 和 #3 以及对照细胞的基因组完整性。未检测到数值或结构异常的证据 (图 1D)。与核转染 (图 1Ei) 和非核转染对照相比,细胞生长和形态正常。与对照 hiPSC 相比,CRISPRa 中的 dCas9 和 tdTomato 表达证实了转基因表达,如 Western blot (补充图 1B,显示克隆 #2 和 #3) 和共聚焦显微镜 (图 1Eii,显示克隆 #2,n = 3 个不同传代) 所示。通过免疫荧光分析干性标记 OCT4 的表达(图 1 Eiii)和流式细胞术分析(显示 94.2% OCT4 和 99.9% TRA1-60 阳性细胞(图 1 Eiv)(显示克隆 #2))来评估多能性。通过在 CRISPRa 和对照系中形成胚状体 (EB) 和定向分化来测试向所有三个胚层的自发分化能力。免疫荧光分析证实了 AFP、β-III-Tu bulin 和 α-平滑肌肌动蛋白 (ACTA2) 的表达,进一步支持内胚层、外胚层和中胚层的命运(图 1 F,显示克隆 #2 和 #3)。转录水平分析表明配对盒 3 ( PAX3 ) 和微管相关蛋白 2 ( MAP2 ) 的表达表明外胚层分化;T-box 转录因子 T ( TBXT ) 表明中胚层命运,而 α-Feto-Protein ( AFP ) 表明内胚层分化(补充图 1 C,显示克隆 #2 和 #3)。我们研究了 CRISPRa 系用于研究通过定向 2D 分化产生的心肌细胞的适用性,这种分化产生了自发跳动的细胞(视频作为补充材料提供),具有强大的 α-辅肌动蛋白 2 (ACTN2) 和心脏肌钙蛋白 T (TNNT2) 心脏标志物表达((补充图 1D,显示为克隆#2)。最后,我们通过确定与心脏肥大和代谢稳态有关的 KLF15 表达的诱导来测试 CRISPRa 系的功能。我们发现,与转染了非靶向 gRNA 的各自亲本系相比,设计用于结合 KLF15 转录起始位点 (TSS) 的 44 bp 5'-上游序列的单个指导 RNA 能够显着增强 CRISPRa 系(克隆#2 和#3)中 KLF15 的转录。对照细胞没有显示独立于转染的 gRNA 的活化(图 1G)。总之,使用完全表征的 hiPSC 系,我们生成了具有纯合靶向插入、正常核型和多能性的人类 CRISPRa 系,并显示出其激活

在什么时机寄生虫攻击并消耗其宿主
图1。NPC的延迟移植可改善势后的长期移植物存活。(a)示意图显示了实验设计。免疫缺陷rag2 - / - 小鼠在1 dpi(急性)或7 dpi(延迟)处局部移植Rfluc表达NPC的局部移植。(b)激光多普勒成像证实中风后脑血流(CBF)减少。(c)中风诱导后2小时对CBF进行定量。(d)代表性的生物发光成像(BLI)说明了两组选定时间点的6周内NPC存活。(e)两组移植后的前3天内对BLI信号的定量。(g)在移植后7天使用EDU掺入的增生评估的示意性时间表,在42天(急性)和35天(延迟)移植后移植时进行染色,以跟踪移植物增殖。(h)在移植后7天,在35 dpi(延迟)和42 dpi(急性)天以35 dpi(延迟)和42 dpi(急性)天的7天和KI67 + NPC对EDU + NPC进行定量的代表性免疫荧光图像。(j)显示具有多能标记Nanog,NPC标记PAX6,Neuronal标记NEUN和星形胶质细胞标记GFAP的表型面板。(k)移植后六周移植的NPC(HUNU+)的代表性免疫荧光图像。比例尺:50µm。(l)急性移植组中移植物组成的定量。数据显示为平均分布,其中红点表示平均值。框图表示数据的25%至75%四分位数。总共使用了8只动物,每组4只动物。箱形图:图中的每个点代表一种动物。线图被绘制为平均值±SEM。使用未配对的Mann-Whitney U检验(C和E)或未配对的t检验(I)评估平均差异的显着性。统计显着性设置为 *,p <0.05; **,p <0.01; ***,p <0.001。

社论:使用计算建模改善干细胞移植递送
干细胞移植已成为再生医学的基石,因为它能够分化为各种细胞类型及其在免疫调节,治疗免疫学疾病和血液学恶性肿瘤中的潜在应用(1)。在各种干细胞类型中,多能胚胎干细胞(ESC)和多能干细胞(ASC)的分化潜力进行了广泛的研究。ESC具有较高的多能性,使它们能够在人体中产生任何细胞类型。然而,围绕其使用的伦理问题导致人们更加关注替代来源,例如诱导的多能干细胞(IPSC)和ASC,包括间质干细胞(MSC),神经干细胞(NSC)和血肿干细胞(HSC)。MSC通过调节T,B,天然杀伤(NK)和树突状细胞来显示免疫调节作用,使其成为自身免疫和炎症性疾病的有前途的工具(2,3)。来自人类脐带血的HSC已广泛用于造血和免疫相关疾病的移植疗法中(4)。HSC移植(HSCT)取得成功,取决于归宿,迁移,植入,自我更新和分化。这些复杂的过程受生长因子,细胞因子和利基相互作用的调节。尽管HSCT具有治疗潜力,但诸如移植物抗宿主病(GVHD),移植排斥和可变的患者结局等挑战持续存在。正在探索诸如免疫耐受性诱导和遗传的策略以及治疗修饰,以增强干细胞的存活和整合(5-8)。正在探索诸如免疫耐受性诱导和遗传的策略以及治疗修饰,以增强干细胞的存活和整合(5-8)。最近的进步表明,将计算模型与免疫数据集成为改善干细胞移植的新途径(9)。机器学习模型可以鉴定重新生成医学中涉及自我更新和谱系规范的关键转录因子和基因网络(10,11)。这些方法还促进了健康干细胞和癌症干细胞(CSC)的比较,这有助于开发恶性肿瘤的靶向疗法(12,13)。免疫学研究主题的前沿,“使用计算建模改善干细胞移植交付”典范这种跨学科方法,并在一系列编译的文章中汇集了开创性的研究,从而贡献了独特的