XiaoMi-AI文件搜索系统
World File Search Systemreplica

超越整体:婴儿宇宙、时空虫洞以及黑洞信息的有序与无序
摘要:20 世纪 80 年代,Coleman 以及 Giddings 和 Strominger 的研究将时空虫洞的物理学与“婴儿宇宙”和一系列理论联系起来。我们重新审视这些想法,使用与负宇宙常数和渐近 AdS 边界相关的特征来强化结果,引入视角的变化,并与最近关于 Page 曲线的复制虫洞讨论联系起来。一个关键的新功能是强调零状态的作用。我们在简单的体拓扑模型中详细探索了这种结构,这些模型使我们能够计算相关边界理论的全部范围。渐近 AdS 希尔伯特空间的维度变成了一个随机变量 Z ,其值可以小于理论中独立状态的简单数量 k 。对于 k > Z ,一致性源于引力路径积分定义的内积的精确退化,因此许多先验独立状态仅相差一个零状态。我们认为,任何一致的引力路径积分都必须具有类似的特性。我们还评论了外推到更复杂模型的其他方面,以及对上述集合中各个成员的黑洞信息问题的可能影响。

引入了用于体内生物传感的全墨捷印刷微针
微针首先是由硅制成的,因为微电子工业为制造综合电路提供了工具,可以适用于微针制造,而硅仍然是最常见的微针材料20。但是,基于洁净室的制造需要复杂的操作和高昂的成本才能实现大规模生产。此外,硅具有可穿戴应用的几个缺点,这就是为什么已经研究了用于微针制造的聚合物材料,金属和其他材料(例如陶瓷)的原因。对于聚合物的微针,越来越明显的是,用于开发下一代聚合物微针的偏爱制造方法和药物输送贴片将是光刻,复制品成型,3D打印和微机械工具20。对于金属微针,光化学蚀刻,电镀和激光切割是最常见的制造技术20。不幸的是,从制造的角度来看,金属微针的制造具有诸如电镀和升降之类的复杂性,这对于质量生产20是不希望的。其他用于微针制造的制造工艺包括注射成型,湿化学蚀刻,反应性离子蚀刻,热压花,激光钻孔,光刻和电型,绘画光刻,两光子聚合和3D打印20。

数字孪生:支持预防性研究的框架...
摘要:由于能够保持遗址的重要性,预防性保护策略逐渐成为比治疗性方法更受青睐的保护策略。此外,现在大多数专家都同意,基于对遗址文化价值的共同理解对遗址进行保护管理对于解决遗址环境的所有特殊性至关重要。最近,大量研究表明,遗产建筑信息模型 (HBIM) 在保护项目中具有协同数据管理的潜力。HBIM 网络平台的最新发展说明了加强数字模型与遗产资产物理领域之间联系的价值。本文提倡应用数字孪生 (DT) 原则,使用 HBIM 模型作为数字复制品,以支持遗址的预防性保护。基于广泛的文献综述,提出了一个将 DT 整合到建筑遗产预防性保护管理计划流程中的综合框架。最后讨论了实施该框架的几项建议,例如识别重要的有形特征、与其完整性相关的威胁以及相应的缓解策略,特别强调价值评估过程。结果是构建预防性保护策略信息的数据模型。该框架提供

随机量子电路中的测量诱导临界性
我们研究了 (Haar) 随机幺正量子电路中投影测量引起的纠缠跃迁的临界行为。使用复制方法,我们将此类电路中纠缠熵的计算映射到二维统计力学模型上。在这种语言中,面积到体积定律纠缠跃迁可以解释为统计力学模型中的有序跃迁。我们使用共形不变性推导出跃迁附近的纠缠熵和互信息的一般缩放特性。我们详细分析了统计力学模型映射到渗流的无限现场希尔伯特空间维度的极限。具体来说,我们使用描述二维渗流临界理论的共形场论的相对较新的结果,计算了子系统大小对数在 n ≥ 1 的 n 次 R'enyi 熵中的普适系数的精确值,并讨论了如何从这个极限访问有限现场希尔伯特空间维度的通用转换,这与二维渗流属于不同的普适性类。我们还评论了与先前在参考文献 1 中研究过的随机张量网络中纠缠转换的关系。
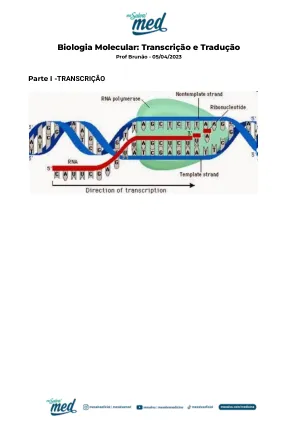
05/04/2023 |分子生物学:转录和翻译。多克克斯
4。(Enem 2011)如今,我们可以说,几乎所有人类都听过DNA及其在大多数生物的遗传中的作用。但是,直到1952年,沃森和克里克的双螺旋桨DNA模型描述的前一年,毫无疑问,这是DNA是遗传物质。在Watson和Crick描述DNA分子的文章中,他们提出了该分子应如何复制的模型。在1958年,梅塞尔森(Meselson)和斯塔尔(Stahl)使用沉重的氮同位素进行了实验,这些同位素被纳入氮基碱基,以评估分子复制的发生方式。从结果来看,他们证实了沃森和克里克所建议的模型,沃森和克里克的基本前提是氮碱基之间的氢桥的破裂。

已发布版本的引文 (APA):Thorsager, M.、Leyva-Mayorga, I.、Soret, B. 和 Popovski, P. (2024)。人工智能通信系统的生成网络层。IEEE 网络快报,6 (2),82-86。文章 10399967。https://doi.org/10.1109/LNET.2024.3354114
如果可以预测,就不需要传达。这是香农将信息定义为不确定性度量的直接结果。然而,数字通信系统主要在源头创建的数据在目的地是不可预测的前提下运行。因此,网络的标准目标是充当比特的哑管道,并确保源数据包的副本通过目的地到达,可能经过多跳。从今以后,中间节点和边缘节点的经典角色是将数据包从输入链路复制到一个或多个输出链路。网络编码 [1] 推广了这一角色,其中网络节点可以超越复制并以更通用的方式组合多个数据流。预测可以显著提高网络层的性能和资源利用率。例如,缓存 [2] 依赖于对可能与目的地相关的数据的预测:源先发制人地将数据传输到边缘节点,该节点存储该数据,直到目的地请求。在这里,源数据不需要实时传输;但边缘节点和目的地都会收到源生成的数据包的副本。网络预测的下一个飞跃是利用生成式人工智能 (GenAI) 及其创建合成数据的能力。具体来说,我们建议使用中间和边缘
突触前cAMP-PKA介导的增强作用诱导海马苔藓纤维末梢处突触囊泡池的重构和通道囊泡的耦合
人们普遍认为,神经回路中的信息存储涉及突触处的纳米级结构变化,从而导致突触印迹的形成。然而,这一假设缺乏直接证据。为了验证这一猜想,我们结合了化学增强、成对突触前后记录的功能分析以及电子显微镜 (EM) 和冷冻断裂复制标记 (FRL) 的结构分析,研究了啮齿动物海马苔藓纤维突触,这是海马三突触回路中的关键突触。突触传递的生物物理分析表明,福斯高林诱导的化学增强分别使易释放囊泡池大小和囊泡释放概率增加了 146% 和 49%。通过 EM 和 FRL 对苔藓纤维突触进行结构分析,发现靠近质膜的囊泡数量和启动蛋白 Munc13-1 簇的数量有所增加,这表明对接囊泡和启动囊泡的数量均有所增加。此外,FRL 分析显示 Munc13-1 和 Ca V 2.1 Ca 2+ 通道之间的距离显著缩短,表明通道-囊泡耦合纳米拓扑结构发生了重构。我们的结果表明,突触前可塑性与活性区的结构重组有关。我们提出,突触囊泡释放位点的潜在纳米组织变化可能与可塑性中枢突触的学习和记忆有关。
![arXiv:2207.01141v5 [quant-ph] 2023 年 4 月 14 日](/simg/8\8c8c25845fd86298065cd7ce5d12178a698e3819.webp)
arXiv:2207.01141v5 [quant-ph] 2023 年 4 月 14 日
在本研究中,我们首先收集并概括了几个现有的非微扰模型,用于描述任意弯曲时空中单个两级量子比特探测器与相对论量子标量场之间的相互作用,其中时间演化由简单生成的幺正体给出,即由施密特秩 1 相互作用哈密顿量生成的幺正体。然后,我们扩展了与这些非微扰模型相关的相对论量子通道,以包括量子场的非常大的一类高斯态,其中包括场上的相干和压缩操作(即高斯操作)的任意组合。我们表明,所有涉及非真空高斯态的物理结果都可以用与真空态相互作用的形式重新表述,但高斯算子通过伴随通道应用于场算子,从而有效地给出了时空中因果传播子形式的高斯运算的“傅里叶变换”解释。此外,我们表明,在这些非微扰模型中,可以精确计算 Rényi 熵,因此,通过复制技巧,可以计算与探测器相互作用后场态的冯·诺依曼熵,而无需对探测器和场的联合初始状态的纯度做出任何假设。这为我们提供了场的三参数“广义猫态”系列,其熵是有限的,并且精确可计算。
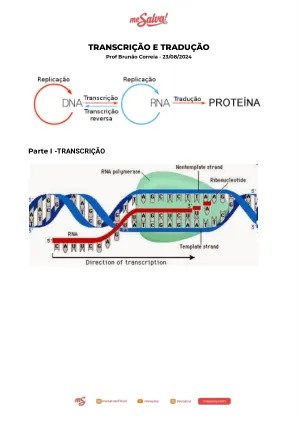
生物学2024 | Brunão|笔录和翻译
4。(Enem 2011)如今,我们可以说,几乎所有人类都听过DNA及其在大多数生物的遗传中的作用。但是,直到1952年,沃森和克里克的双螺旋桨DNA模型描述的前一年,毫无疑问,这是DNA是遗传物质。在Watson和Crick描述DNA分子的文章中,他们提出了该分子应如何复制的模型。在1958年,梅塞尔森(Meselson)和斯塔尔(Stahl)使用沉重的氮同位素进行了实验,这些同位素被纳入氮基碱基,以评估分子复制的发生方式。从结果来看,他们证实了沃森和克里克所建议的模型,沃森和克里克的基本前提是氮碱基之间的氢桥的破裂。

使用无人机和强化学习技术自动进行飞机扫描检查
在飞机日常维护中,通常需要对飞机外表面进行目视检查。使用配备传感器的移动机器人进行自动检查以替代耗时且容易出错的手动检查已成为一种趋势。利用机器学习和数据科学的力量,可以使用图像和点云等感测数据进一步表征缺陷。在这种机器人检查过程中,需要精确的飞机数字模型来规划检查路径,然而,飞机维修店通常无法获得飞机的原始 CAD 模型。因此,使用诸如 3D 激光扫描仪和 RGB-D(红、绿、蓝和深度)相机等传感器,因为它们能够以有效的方式生成感兴趣对象的 3D 模型。本文介绍了一种两阶段方法,使用配备 RGB-D 相机的 UAV(无人驾驶飞行器)自动扫描飞机,以便在无法获得飞机原始 CAD 模型的情况下重建飞机的数字复制品。在第一阶段,无人机相机系统遵循预定义的路径快速扫描飞机并生成飞机的粗略模型。然后,根据飞机的粗略模型计算全覆盖扫描路径。在第二阶段,无人机相机系统遵循计算出的路径对飞机进行紧密扫描,以生成飞机的密集而精确的模型。我们解决了飞机的覆盖路径规划 (CPP) 问题