XiaoMi-AI文件搜索系统
World File Search Systemreplica



提高性能的算法设计... - DergiPark
• 发送日期 / 收到日期:2018 年 7 月 12 日 • 修改发送日期 / 收到修订版:2018 年 8 月 29 日 • 喀布尔日期 / 接受日期:2018 年 9 月 17 日 摘要 提出了一种改进的基于软件的复制相关器方法,该方法适用于微处理器控制的声纳浮标系统,可提高水下物体的探测性能。在海洋中,微处理器控制的浮标可自动进行无线扫描,以清除水下信息,并将其无线传输到主控制站进行额外处理和最终决策。本研究介绍了具有 7-31 单元复制相关的声纳浮标性能和系统设计方案。虽然数字延迟线用于最小化基于硬件的副本相关器的复杂性,但微处理器控制的浮标内所提出的软件副本相关系统已经提高了性能。关键词:浮标、微处理器控制、复制相关、声纳、水下监视。Denize veya okyanus içerisinde、mikroişlemci kontrollü şamandıra、sualtı bilgisinin çıkarılması için otomatik olarak taranması ve daha ileri işlem ve nihai kontrol eylemi sağlanması amacıyla bir uzak yer istasyonuna kablosuz olarak iletilmesi sağlanır。Bu çalışmada,7-31 hücre Replica korelasyonunu içeren sonar şamandıra Performansı ve sistem tasarım taslağı sunulmuştur。Donanımsal replica korelatörünün karmaşıklığı, dijital gecikme kanalları kullanılarak en aza indirilmesine rağmen, önerilen mikroişlemci kontrollü şamandıra, yazılım aracılığıyla replika korelasyonu gerçekleştirilmiş ve geliştirilmiş sistemle Performans artırılmıştır。Anahtar Kelimeler:Şamandra、mikroişlemci kontroller、replika korolasyon、声纳、sualtı gözetim

参议院第 35 号决议
虽然敖德萨的艾伦·诺(Ellenno˝l)艺术博物馆全年展出世界一流的展览,而米德兰(Midland)的海莉纪念图书馆和历史中心则是第一个挂在阿拉莫(Alamo)的钟声,以及有关牛仔和美国西南部的大量口服和书面档案。位于米德兰(Midland)和敖德萨(Odessa)之间的是瓦格纳(Wagner)表演艺术中心,这是一个最先进的设施,在其剧院和演奏厅之间拥有2,000个席位;参观者和居民都在敖德萨的德克萨斯大学二叠纪盆地校园以及莎士比亚学院莎士比亚的环球剧院复制品的现场作品上都享受巨石般的复制品;米德兰音乐节芭蕾舞团,西德克萨斯交响乐团和屡获殊荣的米德兰社区剧院提供了启发和娱乐的现场表演;和
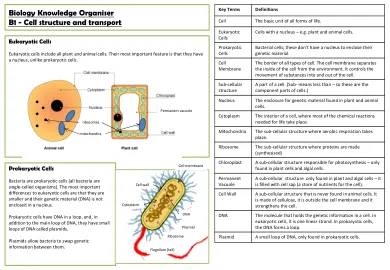
生物学知识组织者B1-细胞结构和运输
细胞在细胞周期中划分以使新细胞,生长和修复。它不如三个单元格分开的简单:必须在执行此操作之前准备。1。细胞生长较大,并产生更多的亚细胞结构,例如核糖体和线粒体。(它为两个单元格制成!)2。通过制作染色体的精确副本,将遗传物质(DNA)加倍。因此,此时有两个副本(在细胞周期图上标记为s)。3。细胞中的微小纤维将每个染色体的副本拉到细胞的相对末端,从而将复制品染色体分开。这意味着细胞核可以分为两个,每个核都带有整个染色体。4。细胞质和细胞膜分裂以形成两个遗传相同的细胞。这是在左图中总结的。

pos(EPS -HEP2023)648-科学论文集
我们提出了一个引人入胜且内容丰富的外展博览会,称为“到目前为止,如此接近”(SFSC),围绕着Alpha磁性光谱仪(AMS)有效载荷操作控制室(AMS POCC)的复制品[1]。这项倡议旨在增强公众对地球各个控制室中进行的对太空任务有效载荷的持续监控和控制的理解。博览会的独特价值在于展示AMS Collaboration的尖端监视软件,从而访问AMS遥测数据。该软件是大流行驱动的创新,允许在限制对CERN站点的物理访问时进行远程参与。POCC的复制品,并由信息丰富的海报和视频补充,是传达基本研究在太空辐射和宇宙射线中重要性的有效工具。

将生物多样性监视数据集成到数字...
目标:DTO-Bioflow将解锁“睡觉”的生物多样性数据,从而使这些和新的生物多样性监测数据的持续流入欧盟数字双海洋。它将为海洋生物学过程的数字复制品创建必不可少的组成部分,将新的和现有的数据流变成基于证据的知识。

一项前瞻性队列研究
麦吉尔大学卫生中心研究所,蒙特利尔,QC,QC,加拿大b麦吉尔大学,蒙特利尔麦吉尔大学医学院,蒙特利尔,加拿大c c c,c,加拿大蒙特利尔大学,c c,加拿大蒙特利尔大学,c c,加拿大蒙特利尔大学,c c,加拿大蒙特利尔大学,c c,加拿大蒙特利尔大学c译院,c c,加拿大蒙特利尔大学,费拉拉·费拉拉大学,意大利法拉拉大学d译本,d 。 Alberta, Canada f Heart and Stroke Strategic Clinical Networks-Alberta Health Services, Alberta, Canada g Department of Internal Medicine III, Division of Endocrinology and Metabolism, Gender Medicine Unit, Medical University of Vienna, Vienna, Austria h Department of Clinical intervention, Science and Technology (CLINTEC), Section for Renal Medicine, Karolinska Institute and Karolinska University hospital, Stockholm, Sweden i Clinical &实验性神经科学(Nice-Imib-iuie),医学院。 默西亚大学,默西亚,西班牙 Canada l Replica Analytics Ltd, Ottawa, Ontario, Canada m Department of Medicine, Internal Medicine, Lausanne University Hospital and University of Lausanne, Lausanne, Switzerland n Center for Psychiatric Epidemiology and Psychopathology, Department of Psychiatry, Lausanne University Hospital and University of Lausanne, Lausanne, Switzerland o Center for Primary Care and Public Health, University of瑞士洛桑(Lausanne)。 Alberta, Canada f Heart and Stroke Strategic Clinical Networks-Alberta Health Services, Alberta, Canada g Department of Internal Medicine III, Division of Endocrinology and Metabolism, Gender Medicine Unit, Medical University of Vienna, Vienna, Austria h Department of Clinical intervention, Science and Technology (CLINTEC), Section for Renal Medicine, Karolinska Institute and Karolinska University hospital, Stockholm, Sweden i Clinical &实验性神经科学(Nice-Imib-iuie),医学院。默西亚大学,默西亚,西班牙Canada l Replica Analytics Ltd, Ottawa, Ontario, Canada m Department of Medicine, Internal Medicine, Lausanne University Hospital and University of Lausanne, Lausanne, Switzerland n Center for Psychiatric Epidemiology and Psychopathology, Department of Psychiatry, Lausanne University Hospital and University of Lausanne, Lausanne, Switzerland o Center for Primary Care and Public Health, University of瑞士洛桑(Lausanne)

黑洞信息之谜与量子德菲内蒂定理
摘要:黑洞信息之谜源于广义相对论与量子理论对黑洞辐射性质的结论存在差异。根据霍金最初的论证,辐射是热的,因此其熵会随着黑洞的蒸发而单调增加。相反,由于量子理论中时间演化的可逆性,辐射熵应该在一定时间后开始减小,正如佩奇曲线所预测的那样。基于复制技巧的新计算证实了这种减小,并揭示了其几何起源:复制品之间形成的时空虫洞。在这里,我们从量子信息论的角度分析了这些结论与霍金最初结论之间的差异,特别是使用了量子德菲内蒂定理。该定理意味着存在额外的信息 W,它既不是黑洞的一部分,也不是辐射的一部分,而是起着参考的作用。通过复制技巧获得的熵可以被识别为以参考 W 为条件的辐射的熵 S ( R | W ),而霍金的原始结果对应于非条件熵 S ( R )。熵 S ( R | W ) 在数学上是集合平均值,在对 N 个独立准备的黑洞进行实验时,它获得了操作意义:对于较大的 N ,它等于它们联合辐射的归一化熵 S ( R 1 · · · RN ) / N 。这个熵和 S ( R ) 之间的差异意味着黑洞是相关的。因此,复制虫洞可以被解释为这种相关性的几何表示。我们的结果还表明广泛使用的随机幺正模型可以扩展到多黑洞,我们通过非平凡检验支持了这一点。