XiaoMi-AI文件搜索系统
World File Search Systemtin

伊利诺伊州HumanaGold®综合Medicare-Medicaid计划预先授权和通知列表,我们更新了我们的预先授权和通知
• Member's Humana ID number, name and date of birth • Date of actual service or hospital admission • Procedure codes, up to 10 maximum per authorization request • Date of proposed procedure, if applicable • Diagnosis codes (primary and secondary), up to 6 maximum per authorization request • Service location • Inpatient location (acute hospital, skilled nursing, hospice) • Outpatient location (telehealth, office, home, off-campus outpatient hospital,校园门诊医院,门诊手术中心[ASC])•转诊(办公室,校外医院,校园内门诊医院,ASC,其他)•税收识别号(TIN)(TIN)和国家提供者标识符(NPI)和服务设施的服务范围•提供服务•服务型•服务型和NPI•服务•服务型•服务•服务•服务范围•服务•服务•服务•服务•服务•服务范围•服务范围•服务范围•服务范围•服务器/npi•相关临床信息•出院计划



第一原理和实验-Repositum
过饱和(Ti,al)的N材料,带有面部的立方(FCC)结构提供了热稳定性和机械性能的独特组合。但是,它们的热诱导的分解过程对于提取其全部潜力至关重要。通过X射线衍射和转移电子显微镜进行了详细的实验研究表明,热力学稳定的Wurtzite型W- ALN的形成以1000 c在100 c的退火温度下(t a)开始,在使用锡(TI,Al)n/tin Multililayerays施加多层式时,以1000 c的形成(t a)。尽管如此,(Ti,al)N/Tin多层的硬度比(Ti,Al)N涂层高100 c t a(900 c),在100 c t a(900 c)下达到32.3±1.0 gpa的峰值,并且硬度下降的趋势随着t的增加而下降。这是因为(ti,al)n分解朝着富含Al和Ti的区域的分层结构,当时与FCC-TIN相干生长。从头算的计算强调,在(Ti,al)N层中的Al优先扩散与锡层相干界面。因此,在一个(ti,al)n层中,更多的层形成,即使在富含质量层的相位变形到w-aln的相变,它们的分层结构仍然存在。一起,计算和实验结果表明,分层的排列具有更高的抵抗力对位错滑行的阻力,并且对涂料完整性是有益的。2022由Elsevier Ltd.这是CC BY-NC-ND许可证(http://creativecommons.org/licenses/by-nc-nd/4.0/)下的开放访问文章。

公众参与报告
1.1.1 2019 年 9 月,土木工程拓展署与规划署联合就新界北第一期发展进行可行性研究,旨在制定新田╱落马洲发展枢纽的初步发展大纲图。新田╱落马洲发展枢纽地区的初步土地用途计划已于 2021 年年中提交立法会。2021 年 10 月,土木工程拓展署与规划署联合开展调查研究,以扩大新田╱落马洲发展枢纽的发展范围,并制定建议发展大纲图,其中考虑到 2021 年施政报告中宣布的北区都会发展策略中有关新田科技城的计划。1.1.2 新田科技城涵盖河套区的香港深圳创新及科技园及新田╱落马洲发展枢纽地区。由于香港深圳创新及科技园已在建设中,建议发展大纲图涵盖河套区以外的地区。 2023 年 5 月 23 日,立法会发展事务委员会听取了有关发展计划的汇报。政府于 2023 年 6 月 6 日至 8 月 5 日(公众参与期)进行了为期两个月的公众参与活动,以收集公众对发展计划的意见。

纪律文件1。程序数据
知识:功能基因组学:对功能基因组原理和应用的理解,包括调节基因活性,基因瘤结构和理解分子生物学的概念。高性能技术:有关转录组学,基因组编辑技术(例如CRISPR/CAS9)和基因组的知识,包括有关功能基因组学中使用的方法及其适用性的知识。多摩学数据集成:了解复杂基因调节网络分析的多摩变数据集成系统和技术的生物学。比较基因组学:关于分子进化,哲学和鉴定物种之间功能元件的观点。疾病中的功能基因组学:了解表型特征和复杂疾病的遗传基础,包括全球基因组关联(GWAS)及其功能性基因组整合和个性化医学。生物信息学:熟悉用于基因组数据分析的生物信息学工具,包括序列比对,基因组障碍和表观基因组数据分析。高级生物信息学工具:有关多摩尼亚数据集成的知识以及智能自动学习在个性化医学的基因组数据分析中的应用。横向技能(角色,专业发展,个人)


新冠疫苗接种服务
皇后大道西134號西營盤賽馬會分科診療所6字樓 6/F, Sai Ying Pun Jockey Club Polyclinic, 134 Queen's Road West 筲箕灣柴灣道8號筲箕灣賽馬會診所地下 G/F, Shau Kei Wan Jockey Club Clinic, 8 Chai Wan Road, Shau Kei Wan 香港仔水塘道10號香港仔賽馬會診所B座地下低層 LG/F, Block B, Aberdeen Jockey Club Clinic, 10 Aberdeen Reservoir Road 灣仔軒尼詩道130號修頓中心二字樓 2/F, Southorn Centre, 130 Hennessy Road, Wan Chai 藍田碧雲道223號藍田社區綜合大樓地下 G/F, Lam Tin Community Complex, 223 Pik Wan Road, Lam Tin 石硤尾南山邨南堯樓地下 G/F, Nam Yiu House, Nam Shan Estate, Shek Kip Mei 大嶼山東涌富東街6號東涌健康中心2座1字樓 1/F, Block 2, Tung Chung Health Centre, 6 Fu Tung Street, Tung Chung, Lantau Island 葵涌葵葉街葵盛東邨盛和樓地下及一樓G/F&1/F,Kwai Yip Street,Kwai Chung,Kwai Yip Street的Shing Wo House,Kwai Chung上水石湖墟马会道108-130 - 1/F,Shek Wu Hui Jockey Club Clinic,108-130骑师俱乐部俱乐部,Sheung Shui shuig Shui将军澳宝琳北路99号将军澳赛马会诊所地下g/f/f/f an o沙田沥源街9号沥源健康院9号沥源健康院33/f,Lek Yuen Health Center,9 Lek Yuen Street,Sha Tin sha Tin荃湾沙咀道113荃湾沙咀道B213 b g/f,B Block B,B街区,Lady Trench Tolench polyclinic,Sha Tsui Road 213卫生中心,塞奇街26号,元长
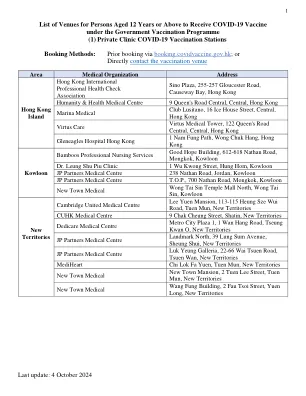
2024 年 10 月 4 日 12 岁以上人士活动场所列表......
北大屿山社区健康中心 (北大屿山医院) 容凤书夫人健康中心 南葵涌赛马会普通科门诊诊所 粉岭家庭医学中心 大埔赛马会普通科门诊诊所 将军澳 (宝宁路) 普通科门诊诊所 天水围 (天业路) 社区健康中心 [疫苗接种服务提供至 2024 年 11 月 18 日。] 伍若瑜夫人普通科门诊诊所* 马鞍山家庭医学诊所* 仁爱普通科门诊诊所* * 疫苗接种服务由 2024 年 11 月 19 日起提供。

新闻通讯
电子束蒸气发生器(EBVG)广泛用于熔化和蒸发金属的应用。由于高度工作温度和真空边界,经常在EBVG腔中熔化和蒸发金属的实验表征变得具有挑战性。计算分析提供了这种物理现象的重要见解。在这项研究中,在三个不同的EBVG腔中研究了TIN的融化和蒸发。这些空腔可以容纳30cc,70cc和110cc的总充电量。内部通用CFD求解器Anupravaha用于CFD模拟融化和蒸发现象。比较了这三个系统的E梁下,熔融池剖面和TIN的蒸发速率的峰值温度。还研究了熔融池表面上的固体氧化物对熔融池轮廓和蒸发速率的影响。观察到,由于对流电流的变化,随着腔体积的增加,蒸发速率的边缘下降。由于纵横比的变化,熔融池的分数增加了70cc和110cc腔。还观察到,由于表面上存在氧化固体,熔融的部分和锡的蒸发速率略有增加。由于固体氧化物层,熔融池轮廓也发生了变化。这种现象可以归因于熔融金属表面上对流电流轮廓的变化。