XiaoMi-AI文件搜索系统
World File Search System切除修复


模板 DNA 链中未修复的碱基切除修复中间体引发复制叉崩溃和 PARP 抑制剂敏感性
DNA 单链断裂 (SSB) 会破坏 DNA 复制并诱导染色体断裂。然而,SSB 存在于复制叉后还是复制叉前时会诱导染色体断裂尚不清楚。为了解决这个问题,我们利用了缺乏 PARP 活性或 XRCC 1 的 SSB 修复缺陷人类细胞对胸苷类似物 5 - 氯-2 0 - 脱氧尿苷 (CldU) 的极佳敏感性。我们表明,在这些细胞中与 CldU 一起孵育会导致染色体断裂、姐妹染色单体交换和细胞毒性,其机制取决于尿嘧啶 DNA 糖基化酶 (UNG) 的 S 期活性。重要的是,我们表明,在一个细胞周期中 CldU 的掺入仅在下一个细胞周期中才具有细胞毒性,此时 CldU 存在于模板 DNA 中。与此一致的是,尽管 UNG 既能诱导复制叉后新生链中的 SSB,也能诱导复制叉前的模板链中的 SSB,但只有后者会触发叉塌陷和染色体断裂。最后,我们表明 BRCA 缺陷细胞对 CldU 高度敏感,无论是单独使用还是与 PARP 抑制剂联合使用,这表明 CldU 可能具有临床实用性。

EPHA5 突变预测...的持久临床益处
TILs:肿瘤浸润淋巴细胞;PFS:无进展生存期;NSCLC:非小细胞肺癌;抗 CTLA-4:抗细胞毒性 T 细胞淋巴细胞-4;PD-L1:程序性死亡配体 1;RTK:Eph 受体酪氨酸激酶;NK:自然杀伤细胞;NGS:靶向下一代测序;DCB:持久临床益处;NDB:无持久益处;OS:总生存期;DFS:无病生存期;GDSC:癌症药物敏感性基因组学;KM:Kaplan-Meier;GO:基因本体论;KEGG:京都基因与基因组百科全书;TCGA:癌症基因组图谱;BER:碱基切除修复;HR:同源重组;MMR:错配修复;FA:范康尼贫血;NER:核苷酸切除修复;NHEJ:非同源末端连接; DSB:DNA 双链断裂;SSB:单链断裂;miRNA:微小RNA;

ASRC-纽约城市大学
星期三,2025 年 1 月 29 日 咖啡 11:30 AM 研讨会 12:00-1:00 PM Riccardo Miggiano 药学副教授 意大利诺瓦拉东皮埃蒙特大学 David Jeruzalmi 化学与生物化学教授 纽约城市大学 细菌核苷酸切除修复途径处理受损 DNA 的机制 摘要 核苷酸切除修复 (NER) 途径是细菌中最重要的 DNA 修复系统之一。UvrABC 核酸外切酶复合物由 UvrA、UvrB 和 UvrC 蛋白组成,构成了负责检测和去除 DNA 损伤的途径。这种多步骤机制需要蛋白质复合物的动态组装,并且依赖于 ATP 结合和水解。具体而言,UvrA 和 UvrB 蛋白对 DNA 进行初步检测以查找损伤,同时避开天然 DNA。

DNA 损伤反应随肌源性分化而下降
DNA 完整性不断面临诱导 DNA 损伤的物质的威胁。所有生物体都配备了 DNA 损伤反应机制网络,可以修复 DNA 损伤并恢复正常的细胞活动。尽管在复制细胞中已经揭示了 DNA 修复机制,但人们对 DNA 损伤在有丝分裂后细胞中的修复方式仍然知之甚少。肌纤维是高度特化的有丝分裂后细胞,以合胞体形式组织,在放射治疗后容易发生与年龄相关的退化和萎缩。我们研究了肌纤维核的 DNA 修复能力,并将其与增殖性成肌细胞中的测量值进行了比较。我们重点研究了纠正电离辐射 (IR) 诱导损伤的 DNA 修复机制,即碱基切除修复、非同源末端连接和同源重组 (HR)。我们发现,在分化程度最高的成肌细胞肌管中,这些 DNA 修复机制表现出 DNA 修复蛋白向 IR 损伤 DNA 募集的动力学减弱。对于碱基切除修复和 HR,这种减弱可能与参与这些过程的关键蛋白的稳态水平降低有关。

环境污染物诱导氧化应激的机制
缩写 8-oxodG 8-氧代-7,8-二氢-2′-脱氧鸟苷 8-oxoGua 8-氧代-7,8-二氢鸟嘌呤 A549 肺泡基底上皮细胞腺癌 AA 花生四烯酸 AhR 芳烃受体 BaP 苯并[a]芘 BEAS-2B 永生化肺上皮细胞 BER 碱基切除修复 CT-DNA 小牛胸腺 DNA CYP 细胞色素 P450 ELISA 酶联免疫吸附试验 EOM 可提取有机物 ETS 环境烟草烟雾 GC/MS 气相色谱/质谱法 HEL 人胚胎肺成纤维细胞 HPLC-MS/MS 高效液相色谱-串联质谱法 IARC 国际癌症研究机构 IsoP 15-F 2t-异前列腺素 IUGR 宫内生长受限 LBW 低出生体重(< 2500 g) LC/GC-MS 液相/气相色谱质谱联用 LPO 脂质过氧化 NER 核苷酸切除修复 NHEJ 非同源末端连接修复 OGG1 8-氧鸟嘌呤 DNA 糖基化酶 PAH 多环芳烃 PBL 外周血淋巴细胞 PGE 2 前列腺素 E2 PM 颗粒物 PTGS 前列腺素内过氧化物合酶 ROS 活性氧 S9 组分 微粒体组分酶 SNP 单核苷酸多态性 UGT UDP-葡萄糖醛酸转移酶 XRCC5 X 射线修复交叉互补 5
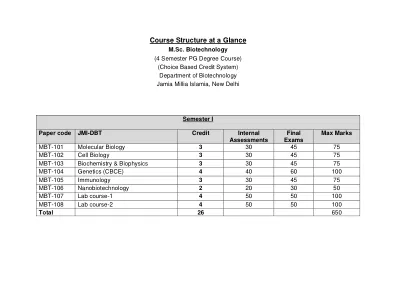
课程结构一览
理学硕士 I 期 MM:75 分子生物学单元 1:基因组的结构和组织(8 小时)染色质组织 - 组蛋白和 DNA 相互作用组、染色质结构、核小体、染色质组织和重塑、染色体、异染色质和真染色质、扭转应力、DNA 拓扑结构 - 链接数、扭曲、扭动、超螺旋、拓扑异构体。第二单元:DNA复制、修复和重组(8 小时)DNA复制模型,Meselson 和 Stahl 实验,DNA聚合酶,病毒、细菌和真核生物中的 DNA 复制,复制叉,复制的校对和保真度,末端复制问题和端粒酶,复制抑制药物,DNA损伤剂,DNA修复机制(核苷酸切除修复、碱基切除修复、错配修复、重组修复、双链断裂修复、转录偶联修复、重组——同源、非同源和位点特异性重组)第三单元:基因表达和调控(8 小时)原核和真核基因的结构、调控区域、转录因子、转录机制、RNA聚合酶、RNA加工结构和不同 RNA 类型的功能、起始复合物的形成、延长、终止;操纵子概念-乳糖操纵子、色氨酸操纵子、arb操纵子、𝜆-阻遏物、lexA阻遏物、噬菌体的溶源性和溶解性循环、核糖开关、转录抑制剂。


软组织肉瘤中核苷酸 - 分离修复(NER)途径的作用:综述和关注其作为治疗靶标的潜力
核苷酸切除修复(NER)途径涉及三十多个蛋白质 - 蛋白质相互作用,并去除化学疗法药物引起的DNA加合物。NER的关键基因通常在癌细胞中过度表达,该途径的改变负责增加或降低对特定治疗剂的敏感性。这在软组织肉瘤(STS)中特别相关,稀有间充质原始肿瘤的潜在机制仍然缺乏理解。完全可以是STS中潜在的治疗靶标。NER活性的微妙调节可能在临床上与替代预后标记或预测对化学疗法剂的敏感性有关。应进一步对NER进行进一步的预期评估以解决这个问题。摘要

骨肉瘤的顺铂耐药性:候选 DNA 修复相关治疗靶点和定制治疗药物的体外验证
高级别骨肉瘤是最常见的骨恶性肿瘤,其治疗主要依靠顺铂和其他 DNA 损伤药物。因此,DNA 修复机制的改变可能会显著影响对化疗的反应或耐药性。在本研究中,我们利用一组对顺铂敏感或耐药的人类骨肉瘤细胞系,评估了属于核苷酸切除修复 (NER) 或碱基切除修复 (BER) 途径的 DNA 修复相关因子以及一组 18 种激酶作为候选治疗靶点的价值,这些激酶在顺铂耐药变体中的表达高于其亲本细胞系,可能间接参与 DNA 修复。通过基因沉默方法和体外逆转 CDDP 耐药性,验证了这些因素与人类骨肉瘤细胞顺铂耐药性的因果关系。这种方法突出了一个基因亚群,蛋白质表达分析进一步证实了它们作为有希望的候选治疗靶点的价值。然后分析了 15 种抑制剂药物针对这些基因或其途径的体外活性,以确定在固有活性和克服顺铂耐药性的能力方面最活跃的药物。NSC130813(NERI02;F06)和雷公藤内酯醇均以 NER 因子为靶点,被证明是两种最活跃的药物,没有证据表明与顺铂有交叉耐药性。联合体外治疗表明,NSC130813 和雷公藤内酯醇与顺铂一起使用时,能够提高其在药物敏感和耐药骨肉瘤细胞中的疗效。这一证据可能表明,对于对顺铂反应性降低的骨肉瘤患者,未来这是一种有趣的治疗选择,即使必须仔细考虑附加附带毒性的可能影响。此外,我们的研究还表明,针对属于丝裂原活化蛋白激酶 (MAPK) 或成纤维细胞生长因子受体 (FGFR) 通路的蛋白激酶可能为骨肉瘤带来新的有希望的治疗前景,需要进一步研究。