XiaoMi-AI文件搜索系统
World File Search System吉兰

在健壮的联合模型设置中利用机器学习
在强大的联合模型设定主管中利用机器学习:Karen Cairns博士和Lisa McFetridge博士自2020年4月以来,贝尔法斯特皇后大学的研究人员通过贝尔法斯特皇家护理部门(ICU)的贝尔法斯特皇后学院的研究人员收集了大量数据。拟议的博士将开发新的统计理论,以更好地说明和分析此类数据的复杂性,并与贝尔法斯特皇家维多利亚医院的呼吸医学顾问紧密合作,计算机科学的高级讲师查尔斯·吉兰(Charles Gillan)将这一理论付诸实践。因此,这是对ICU患者产生现实世界影响的一个令人兴奋的机会。该项目将开发的新方法将用于确定严重不良事件的风险因素,并产生动态预测,这些预测将被纳入预警系统中以帮助ICU员工。机械通气是一种挽救生命的疗法,但不幸的是,它确实对患者构成风险。预防策略的早期鉴定和发展对于最终改善需要机械通气的患者的生存预后至关重要。该项目将开发新的鲁棒关节建模方法来同时分析ICU患者在住院期间从ICU患者收集的重复测量(纵向数据),并且这些生物标志物的动态变化对患者预后的影响(McFetridge 2021)。生存森林是一种合奏技术,类似于随机森林(Breiman 2001),它在事实上的环境中应用(Ishwaran 2008)。尤其是,这项工作将通过掺入生存树和随机生存森林来利用机器学习技术的准确性,以更好地捕获活动时间的过程。与标准生存模型(例如COX回归)相比,它们已显示出高度准确的集合预测指标,提高了预测精度。因此,它们在健壮的关节模型设置中的利用将提供更精确的动态个性化的生存预测,同时考虑了生物标志物随时间的发展,并且有可能存在趋势不符合人群平均值的外围个体的共同存在的潜力。这种强大的方法将在ICU中为严重的不良事件奠定急需的预警系统的基础。该博士将融入针对预警系统开发和完善的更广泛的项目集合,学生与整个研究团队紧密合作,以实施所需的现实世界影响。有关该项目的更多详细信息,请联系主要主管(k.cairns@qub.ac.uk)。McFetridge,L.M.,Asar,Ö。和沃林(J. 63(8),1587-1606。 Breiman,L。随机森林,机器学习,2001年; 45,5-32。McFetridge,L.M.,Asar,Ö。和沃林(J. 63(8),1587-1606。Breiman,L。随机森林,机器学习,2001年; 45,5-32。

* 通讯作者:Mohammad-Amin Abshat (MD),伊朗拉什特吉兰医科大学医学院。Seyed Javad Masoumi (MSc),伊朗德黑兰大学心理学与教育学院心理学系。电子邮件:aminabshat9777@gmail.com 电子邮件:masoumi.seyedjavad@gmail.com 如何引用本文:Khani A、Sadeghi Velni E、Hosseini SS、Bagherieh D、Abshat MA、Masoumi SJ。人工智能:一种改善重症监护病房患者护理的宝贵而有效的工具。J Nurs Rep Clin Pract。2024 年。
AI 在 ICU 护理中最重要的贡献之一是它能够实时处理大量患者数据 [5]。ICU 患者通常使用各种设备进行监测,这些设备可跟踪生命体征,包括心率、血压、血氧饱和度和呼吸模式。AI 算法可以即时分析这些数据,识别模式并在并发症变得危急之前预测它们。例如,AI 可以通过识别患者生理数据的细微变化来检测败血症(一种危及生命的疾病)的早期迹象。这种预测能力使护士能够尽早进行干预,从而有可能预防严重的并发症并提高患者的存活率 [5]。

COVID-19 疫苗犹豫及其驱动因素
2020 年 3 月 11 日,世界卫生组织 (WHO) 宣布冠状病毒病 (COVID-19) 为全球大流行病。此后,COVID-19 大流行造成了前所未有的公共卫生威胁,时隔两年仍在全球范围内持续存在。虽然一些国家已转入地方性流行阶段,但世卫组织警告称,大流行远未结束,如果疫苗接种水平下降,可能继续引发全球疫情 [1,2]。自 2020 年底新冠疫苗获批以来,疫苗接种一直是各国实现群体免疫的重点 [3,4]。从公共卫生角度来看,大规模接种疫苗被视为抗击传染病爆发最有效的措施 [5-7]。截至 2022 年 10 月 25 日,马来西亚已报告近 490 万例新冠病例,总人口的 84.3% 接种了新冠疫苗。按照全球标准,马来西亚的疫苗接种率与其他国家相比相对较高。尽管如此,随着马来西亚向地方性流行阶段过渡,疫苗犹豫(世卫组织将其定义为“尽管有疫苗接种服务,但仍延迟接受或拒绝接种疫苗” [8])仍然是部分民众面临的挑战 [2,9]。例如,截至 2022 年 10 月 25 日,80 岁以上的人群(80.9%)和马来西亚 16 个州中的 6 个州(吉打(77.4%)、吉兰丹(65.0%)、彭亨(76.0%)、霹雳(80.7%)、沙巴(68.2%)、登嘉楼(73.4%))的初级疫苗接种覆盖率低于全国平均水平 [10]。尽管马来西亚的整体疫苗接种覆盖率相对较高,但疫苗犹豫仍然是一个值得关注的问题,主要有三个原因。首先,未接种疫苗者的死亡率很高。截至 2022 年 10 月 25 日,在该国记录的 36,452 例 COVID-19 死亡病例中,61.2% 未接种疫苗 [10]。其次,提高疫苗接种覆盖率不仅会影响死亡率,而且还与积极的公共和个人健康结果相关,例如降低 COVID-19 的传播率并减少 COVID-19 感染引起的严重程度和并发症 [11,12]。第三,目前需要疫苗加强剂来增强或恢复在主要疫苗接种后可能随着时间的推移而减弱的保护作用,这危及疫苗覆盖的有效性 [13-15]。因此,至少在可预见的未来,需要继续接种疫苗。因此,获得有关疫苗犹豫态度持续存在的原因的行为洞察对于有效管理大流行至关重要 [16]。鉴于马来西亚卫生部长的评论,这一点尤其重要,他确认,采用行为科学对于对抗击 COVID-19 的后续传播至关重要 [17]。马来西亚于 2021 年 2 月 24 日启动了 COVID-19 疫苗接种计划。但是,要求希望接种疫苗的居民在接种疫苗前进行登记。因此,与马来西亚之前大多数研究调查了整个人口的疫苗接种意向而不考虑其登记状态的研究不同[18-20],这项研究缩小到未登记人口群体的疫苗接种意向,这代表了

里海沿岸国家的环境研究
生物学系,科学系,吉兰大学,南乔街,P.O.Box 1914,伊朗,伊朗,电话:0098-9113330017,传真:0098-131-3233647,电子邮件:umistbiology20@gmail.com; salehiz@guilan.ac.ir里海是世界上最大的陆上水域。它是地球上最大的封闭水体(Roshan等人。2012),占湖泊水域全球量的44%。与世界上其他半封闭和封闭的海洋相比,对里海的可变性知之甚少(Ibrayev等人。2010)。里海海洋受到环境威胁的巨大压力,例如海水水平的变化,捕捞过多,风险前锋海洋,侵扰工业和农业以及发展大多数中海国家的城市(Karrari等人)(Karrari等人2012; Jamalomidi 2013)。里海是一个封闭的水体,在中亚地区起着重要的地缘政治作用。在过去的几十年中,自然和人为因素的联合作用一直在加剧里海中的环境状态。不断增加的人类活动,例如石油和天然气行业,特别是在里海,渔业,农业和旅游业的北部,以及数十年的环境管理不善,导致了水质的严重退化(Fathabadi等人。2012; Fendereski 2014)。里海中最典型的有毒物质是石油烃,重金属,苯酚,表面活性剂和氯 - 有机农药(Aladin&Plotnikov 2004)。由于人为污染,它面临着重大的环境挑战。地理位置和碳氢化合物资源的存在(石油和天然气)使里海地区对沿海国家和主要世界大国的地球缘缘地区至关重要。里海地区的几个州包括里海的五个沿海国家:伊朗伊斯兰共和国,土库曼斯坦,哈萨克斯坦,俄罗斯联邦和阿塞拜疆共和国。里海环境面临的主要挑战包括水位上升,环境污染,外来物种进入里海海的入口,植物园的丧失和富营养化。分析性描述性研究试图回答这个基本问题:“里海沿岸国家对环境损害有什么责任?”可以说,里海的沿海国家单独或集体负责其自身的遗漏和行动,从而造成环境破坏。因为在里海及其沿海国家的各个方面的研究很重要,所以我们很高兴认识到为环境和农业研究做出贡献的研究人员的努力。本期特刊中的研究为生物,生态和农业研究提供了广泛的看法,该研究应为环境研究的未来研究提供信息和启发。Toshbekov等。(2024)在“北极狐狸的行为适应,vulpes lagopus响应气候变化”中促成了这一问题。Umirzokov等。(2024)在“里海沉积物的生化分析:对环境污染和生物修复的影响”中探讨了这个问题。他们的研究调查了北极狐狸,紫罗护拉戈普斯的行为适应,以应对气候变化,重点是阿拉斯加北部的三年(2021-2023)的狩猎模式的变化,DEN场地选择和社交互动的变化,并雇用了GPS追踪60 Foxes的GPS,100 Fox,100个远程相机陷阱和直接的现场观察。他们为里海沉积物的污染状况和微生物生态学提供了全面的见解,揭示了内在生物修复的巨大污染和显着的潜力。

全球化与发展 - 拉美和加勒比经济委员会资料库
本文件的编写工作由拉加经委会执行秘书何塞·安东尼奥·奥坎波协调,并与拉加经委会阿根廷办事处前主任胡安·马丁合作; Reynaldo Bajraj,副执行秘书; Alicia Bárcena,可持续发展和人类住区司司长;芭芭拉·斯托林斯 (Barbara Stallings),经济发展司司长; Vivianne Ventura-Dias,国际贸易与一体化司司长;尤金尼奥·拉赫拉,顾问;以及委员会秘书特别助理 María Elisa Bernal。Jean Acquatella、Oscar Altimir、Mario Cimolli、Ricardo Ffrench-Davis、Len Ishmael、Luis Felipe Jiménez、Jorge Katz、Manuel Marfán、Jorge Martínez、Juan Carlos Ramírez、Daniel Titelman、Andras Uthoff 和 Miguel Villa 参与了编写个别章节。以下人员参与了其内容的制定和讨论:María José Acosta、Hugo Altomonte、José Pablo Arellano、Irma Arriagada、Carmen Artigas、Renato Baumann、Rudolf Buitelaar、Inés Bustillo、Alvaro Calderón、Berverley Carlson、Jessica Cuadros 、卡洛斯·德米格尔、马丁·迪尔文、埃尔南多帕索、何塞·埃利亚斯·杜兰、休伯特·艾斯凯斯、埃内斯托·埃斯平多拉、费利佩·费雷拉、胡安·卡洛斯·费雷斯、吉尔伯托·加洛平、弗朗西斯科·加托、克里斯蒂安·吉默斯、何塞·哈维尔·戈麦斯、丽贝卡·格林斯潘、约翰内斯·赫尔曼、迈克尔·亨德里克森、伊莎贝尔·埃尔南德斯、马丁·霍费尔特、安德烈·霍费尔特、马丁·霍本海恩,里卡多·乔丹、桑山干雄、阿图罗·莱昂、胡安·卡洛斯·莱尔达、卡拉·马卡里奥、海伦·麦克贝恩、里卡多·马特纳、豪尔赫·马塔尔、何塞·卡洛斯·席尔瓦·马托斯、格雷谢拉·莫吉兰斯基、索尼娅·蒙塔尼奥、塞萨尔·莫拉莱斯、胡安·卡洛斯·莫雷诺·布里德、迈克尔·莫蒂莫尔、吉列尔莫、Mun乔治娜·努涅斯、玛丽亚·安吉拉·帕拉、威尔逊佩雷斯、埃斯特万·佩雷斯、玛丽安·沙佩尔、伊万·席尔瓦、维罗妮卡·席尔瓦、安娜·索霍、安德烈斯·索利马诺、罗杰里奥·斯图达特、乔瓦尼·斯图姆波、拉奎尔·萨拉赫曼、特鲁迪·蒂鲁克辛、伊恩·汤姆森、赫尔维亚·维洛索、于尔根·韦勒和里卡多·萨帕塔。Armando Di Fillipo、Edmund V. FitzGerald、Wolf Grabendorf、Stephany Griffith-Jones、Gabriel Palma 和 Santiago Perry 担任顾问。

通过RNA识别的先天免疫反应 - 生物化学
Tollike受体:对先天免疫的最新见解和观点。免疫,57,649 - 673。4)Rehwinkel,J。&Gack,M.U。(2020)RIG-I样受体:它们在RNA传感中的调节和作用。nat。修订版免疫。,20,537 - 551。5)Venkataraman,T.,Valdes,M.,Elsby,R.,Kakuta,S.,Cace- Res,G.,Saijo,S.,Iwakura,Y。,&Barber,G.N。(2007)DEXD/H盒RNA解旋酶LGP2的损失表现出不同的抗病毒反应。J. Immunol。 ,178,6444 - 6455。 6)Satoh,T.,Kato,H.,Kumagai,Y.,Yoneyama,M.,Sato,S.,Matsushita,K.,Tsujimura,T.,Fujita,T. (2010)LGP2是RIG-II和MDA5介导的抗病毒反应的积极调节剂。 proc。 natl。 学院。 SCI。 美国,107,1512 - 1517。 7)Bruns,A.M. (2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。 mol。 单元格,55,771 - 781。 8)乌鸦,Y.J. &Stetson,D.B。 (2022)I型干扰素:10年了。 nat。 修订版 免疫。 ,22,471 - 483。 9)村上,S。 (2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。 单元格,82,2236 - 2251。 10)Ablasser,A。 &Chen,Z.J。 (2019)CGA在行动中:在免疫和炎症中扩大角色。 Science,363,EAAT8657。J. Immunol。,178,6444 - 6455。6)Satoh,T.,Kato,H.,Kumagai,Y.,Yoneyama,M.,Sato,S.,Matsushita,K.,Tsujimura,T.,Fujita,T.(2010)LGP2是RIG-II和MDA5介导的抗病毒反应的积极调节剂。proc。natl。学院。SCI。 美国,107,1512 - 1517。 7)Bruns,A.M. (2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。 mol。 单元格,55,771 - 781。 8)乌鸦,Y.J. &Stetson,D.B。 (2022)I型干扰素:10年了。 nat。 修订版 免疫。 ,22,471 - 483。 9)村上,S。 (2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。 单元格,82,2236 - 2251。 10)Ablasser,A。 &Chen,Z.J。 (2019)CGA在行动中:在免疫和炎症中扩大角色。 Science,363,EAAT8657。SCI。美国,107,1512 - 1517。7)Bruns,A.M.(2014)先天免疫传感器LGP2通过调节MDA5 - RNA相互作用和弹性组件来激活抗病毒信号传导。mol。单元格,55,771 - 781。8)乌鸦,Y.J.&Stetson,D.B。(2022)I型干扰素:10年了。nat。修订版免疫。,22,471 - 483。9)村上,S。(2022)mRNA中的隐藏代码:m(6)a对基因表达的控制。摩尔。单元格,82,2236 - 2251。10)Ablasser,A。&Chen,Z.J。(2019)CGA在行动中:在免疫和炎症中扩大角色。Science,363,EAAT8657。11)Ablasser,A。&Hur,S。(2020)调节CGAS和RLR介导的对核酸的免疫力。nat。免疫。,21,17 - 29。12)Hopfner,K.P。&Hornung,V。(2020)CGAS刺信信号传导的分子机制和细胞功能。nat。修订版mol。细胞生物。 ,21,501 - 521。 13)伦纳德(J.N.),吉兰多(R. (2008)TLR3通过合作受体二聚体信号形式。 proc。 natl。 学院。 SCI。 美国,105,258 - 263。 14) (2008)带有双链RNA的Toll样重复3信号传导的结构基础。 Science,320,379 - 381。 15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。 (2005)Toll样受体3配体结合结构域的分子结构。 proc。 natl。 学院。 SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。细胞生物。,21,501 - 521。13)伦纳德(J.N.),吉兰多(R.(2008)TLR3通过合作受体二聚体信号形式。proc。natl。学院。SCI。 美国,105,258 - 263。 14) (2008)带有双链RNA的Toll样重复3信号传导的结构基础。 Science,320,379 - 381。 15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。 (2005)Toll样受体3配体结合结构域的分子结构。 proc。 natl。 学院。 SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。SCI。美国,105,258 - 263。14)(2008)带有双链RNA的Toll样重复3信号传导的结构基础。Science,320,379 - 381。15)Bell,J.K.,Botos,I.,Hall,P.R。,Askins,J.,Shiloach,J.,Segal,D.M。和Davies,D.R。(2005)Toll样受体3配体结合结构域的分子结构。proc。natl。学院。SCI。 美国,102,10976 - 10980。 16)Choe,J.,Kelker,M.S。和Wilson,I.A。 (2005)人Toll样受体3(TLR3)外生域的晶体结构。 科学,309,581 - 585。 17)塔布塔(K. (2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。 proc。 SCI。SCI。美国,102,10976 - 10980。16)Choe,J.,Kelker,M.S。和Wilson,I.A。(2005)人Toll样受体3(TLR3)外生域的晶体结构。科学,309,581 - 585。17)塔布塔(K.(2004)Toll样受体9和3作为对小鼠巨细胞病毒感染的先天免疫防御的重要组成部分。proc。SCI。SCI。natl。学院。美国,101,3516 - 3521。18)Davey,G.M.,Wojtasiak,M.,Proietto,A.I.,Carbone,F.R。,Heath,W.R。,&Bedoui,S。(2010)剪切边缘:CD8 T细胞免疫的启动:Surpes Simperx Simplex Virus 1型需要Cognate Tlr3在Vivo中的表达。J. Immunol。 ,184,2243 - 2246。 19)Oshiumi,H.,Okamoto,M.,Fujii,K.,Kawanishi,T.,Matsu-Moto,M.,Koike,S。,&Seya,T。(2011)TLR3/TICAM-1途径是对Poliovi-Rus Rus Infection的先天免疫反应的强制性。 J. Immunol。 ,187,5320 - 5327。 20)张,S.Y. (2007)疱疹患者的TLR3缺乏效率 -J. Immunol。,184,2243 - 2246。19)Oshiumi,H.,Okamoto,M.,Fujii,K.,Kawanishi,T.,Matsu-Moto,M.,Koike,S。,&Seya,T。(2011)TLR3/TICAM-1途径是对Poliovi-Rus Rus Infection的先天免疫反应的强制性。J. Immunol。 ,187,5320 - 5327。 20)张,S.Y. (2007)疱疹患者的TLR3缺乏效率 -J. Immunol。,187,5320 - 5327。20)张,S.Y.(2007)疱疹患者的TLR3缺乏效率 -
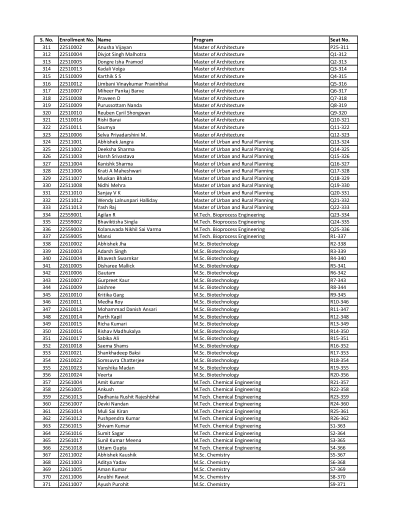
序号 入学编号 姓名 课程 座位号 311 22510002 Anusha Vijayan 建筑学硕士 P25-311 312 22510004 Divjot Singh Malhotra 建筑学硕士
序号 入学编号 姓名 课程 座位号 311 22510002 Anusha Vijayan 建筑学硕士 P25-311 312 22510004 Divjot Singh Malhotra 建筑学硕士 Q1-312 313 22510005 Dongre Isha Pramod 建筑学硕士 Q2-313 314 22510013 Kadali Volga 建筑学硕士 Q3-314 315 21510009 Karthik SS 建筑学硕士 Q4-315 316 22510012 Limbani Vinaykumar Pravinbhai 建筑学硕士 Q5-316 317 22510007 Miheer Pankaj Barve 建筑学硕士 Q6-317 318 22510008 Praveen D 建筑学硕士Q7-318 319 22510009 Purussottam Nanda 建筑学硕士 Q8-319 320 22510010 Reuben Cyril Shongwan 建筑学硕士 Q9-320 321 21510016 Rishi Barai 建筑学硕士 Q10-321 322 22510011 Saumya 建筑学硕士 Q11-322 323 22510006 Selva Priyadarshini M. 建筑学硕士 Q12-323 324 22511001 Abhishek Jangra 城市与乡村规划硕士 Q13-324 325 22511002 Deeksha Sharma 城市与乡村规划硕士 Q14-325 326 22511003 Harsh Srivastava 城市与乡村规划硕士Q15-326 327 22511004 Kanishk Sharma 城市与乡村规划硕士 Q16-327 328 22511006 Krati A Maheshwari 城市与乡村规划硕士 Q17-328 329 22511007 Muskan Bhakta 城市与乡村规划硕士 Q18-329 330 22511008 Nidhi Mehra 城市与乡村规划硕士 Q19-330 331 22511010 Sanjay VK 城市与乡村规划硕士 Q20-331 332 22511012 Wendy Lalnunpari Halliday 城市与乡村规划硕士 Q21-332 333 22511013 Yash Raj 城市与乡村规划硕士 Q22-333 334 22559001阿吉兰·R M.Tech。生物过程工程 Q23-334 335 22559002 Bhaviktisha Singla M.Tech.生物过程工程 Q24-335 336 22559003 Kolanuvada Nikhil Sai Varma M.Tech.生物过程工程 Q25-336 337 22559005 Mansi M.Tech.生物过程工程 R1-337 338 22610002 Abhishek Jha 硕士生物技术 R2-338 339 22610003 Adarsh Singh 硕士生物技术 R3-339 340 22610004 Bhavesh Swarnkar 硕士生物技术 R4-340 341 22610005 Disharee Mallick 硕士生物技术 R5-341 342 22610006 Gautam 硕士生物技术 R6-342 343 22610007 Gurpreet Kaur 硕士生物技术 R7-343 344 22610009 Jaishree 硕士生物技术 R8-344 345 22610010 Kritika Garg 硕士生物技术 R9-345 346 22610011 Medha Roy 硕士生物技术 R10-346 347 22610013 MohammadDanish Ansari 理学硕士生物技术 R11-347 348 22610014 Parth Kapil 硕士生物技术 R12-348 349 22610015 Richa Kumari 硕士生物技术 R13-349 350 22610016 Rishav Madhukalya 硕士生物技术 R14-350 351 22610017 Sabika Ali 硕士生物技术 R15-351 352 22610018 Saema Shams 硕士生物技术 R16-352 353 22610021 Shankhadeep Baksi 硕士生物技术 R17-353 354 22610022 Somsuvra Chatterjee 硕士生物技术 R18-354 355 22610023 Vanshika Madan 硕士生物技术 R19-355 356 22610024 Veerta 硕士生物技术 R20-356 357 22561004 Amit Kumar M.Tech。化学工程 R21-357 358 22561005 Ankush M.Tech。化学工程 R22-358 359 22561013 Dadhania Rushit Rajeshbhai M.Tech.化学工程 R23-359 360 22561007 Devki Nandan M.Tech.化学工程 R24-360 361 22561014 Muli Sai Kiran M.Tech.化学工程 R25-361 362 22561012 Pushpendra Kumar M.Tech.化学工程 R26-362 363 22561015 Shivam Kumar M.Tech.化学工程 S1-363 364 22561016 Sumit Sagar M.Tech.化学工程 S2-364 365 22561017 Sunil Kumar Meena M.Tech.化学工程 S3-365 366 22561018 Uttam Gupta M.Tech.化学工程 S4-366 367 22611002 Abhishek Kaushik 硕士化学 S5-367 368 22611003 Aditya Yadav 硕士化学 S6-368 369 22611005 Aman Kumar 硕士化学 S7-369 370 22611006 Anubhi Rawat M.Sc.化学 S8-370 371 22611007 Ayush Purohit M.Sc.化学 S9-371