XiaoMi-AI文件搜索系统
World File Search System同源

保守的 Gsx2/Ind 同源结构域单体与同源二聚体 DNA 结合决定了果蝇和小鼠的调控结果
1 辛辛那提儿童医院研究基金会分子与发育生物学研究生项目,美国俄亥俄州辛辛那提 45229;2 辛辛那提大学医学院医学科学家培训项目,美国俄亥俄州辛辛那提 45229;3 辛辛那提大学医学院辛辛那提儿童医院医学中心发育生物学科,美国俄亥俄州辛辛那提 45229;4 美国国立卫生研究院国家眼科研究所眼科遗传学和视觉功能分部,美国马里兰州贝塞斯达 20892;5 辛辛那提大学生物医学工程系,美国俄亥俄州辛辛那提 45219;6 辛辛那提大学医学院儿科系,美国俄亥俄州辛辛那提 45229; 7 美国俄亥俄州辛辛那提市辛辛那提儿童医院医学中心生物医学信息学部,辛辛那提 45229;8 美国俄亥俄州辛辛那提市辛辛那提大学医学院生物医学信息学系,辛辛那提 45229
探索种系与同源重组的躯体起源
1汤姆·贝克癌症中心,卡明医学院,加拿大卡尔加里; 2法国维勒维夫大学,巴黎 - 萨克莱大学的Gustave Roussy研究所;日本千叶北部国家癌症中心医院3;澳大利亚墨尔本市彼得·麦卡伦(Peter MacCallum Cancer Center)4; 5号国家癌症中心,大韩民国戈阳; 6奥克兰市医院和奥克兰大学,新西兰奥克兰;西班牙巴塞罗那市Vall d'Hebron肿瘤学研究所(VHIO)Vall D'Hebron大学医院7; 8 Irccs iStituto romagnolo per lo Studio dei tumori(Irst)Dino Amadori,意大利梅尔多拉; 9英国格拉斯哥的苏格兰癌症中心的Beatson,格拉斯哥大学癌症科学学院9;威斯康星州麦迪逊市威斯康星州麦迪逊市威斯康星大学10号; 11 PUCRS医学院,巴西Porto Alegre; 12 Clinique Victor Hugo Center Jean Bernard,法国勒芒; 13德国海德堡海德堡大学医院国家肿瘤疾病中心(NCT);挪威洛伦斯科格(Lørenskog)14 Akershus大学医院(Ahus); 15美国北卡罗来纳州达勒姆市Pfizer Inc.; 16美国宾夕法尼亚州大学维尔市Pfizer Inc.; 17 Pfizer Inc.,美国加利福尼亚州拉霍亚; 18犹他大学,犹他州盐湖城,美国犹他州,亨斯曼癌症研究所(NCI-CCC)

同源重组修复(HRR)的基因检测...
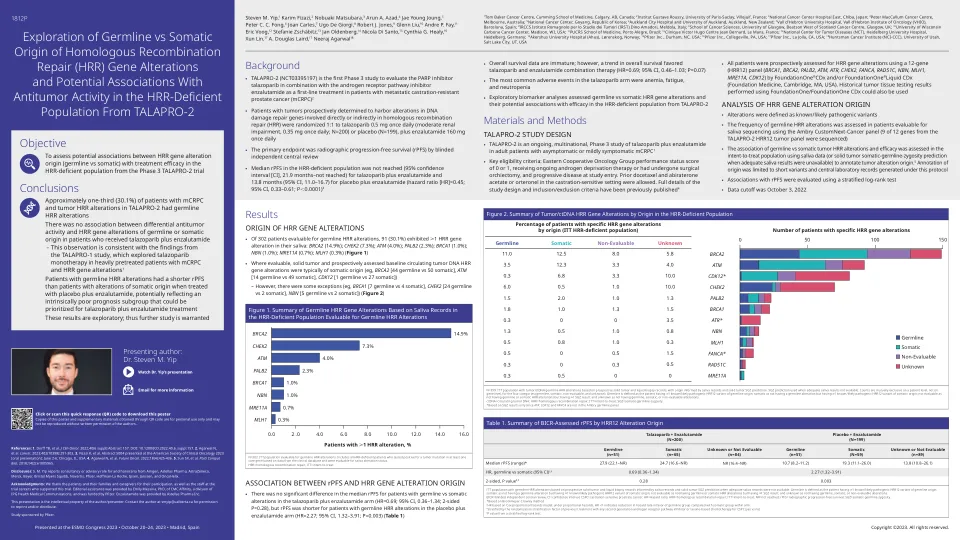
转移性去势抵抗性前列腺癌 (mCRPC) 患者的平均生存期仅为 13 个月。在多达四分之一的 mCRPC 患者中,同源重组修复 (HRR) 通路中发现了新的可预测和可操作的生物标志物,这促使多聚 ADP 核糖聚合酶抑制剂 (PARPi) 等靶向疗法获得批准,有可能改善生存结果。PARPi 的批准促使美国国家综合癌症网络 (NCCN) 等指导机构积极推荐进行种系和/或体细胞 HRR 基因组测试,以确定哪些患者将受益于 PARPi。然而,由于基因检测仍处于早期阶段,尤其是在低收入和中等收入国家,成本和可用性是主要障碍,因此存在一些挑战。此外,还存在一些问题,例如选择最佳组织进行基因检测、存档、储存、检索组织块、解释和分类 HRR 通路中的变异,以及测试前和测试后的遗传咨询的必要性。本综述深入分析了 mCRPC 中普遍存在的 HRR 基因突变以及更广泛的基因检测所面临的挑战,以识别 HRR 通路中可操作的种系致病变异和体细胞突变,并提出了一种临床算法来提高基因检测过程的效率。

果蝇 Myc 直系同源基因模型
(A) 果蝇 (Drosophila melanogaster) 和菠萝蜜 (D. ananassae) 中 Myc 基因组邻域的同源性比较。细箭头表示果蝇 (D. melanogaster) (顶部) 和菠萝蜜 (D. ananassae) (底部) 中目标基因 Myc 所在的 DNA 链。指向右侧的细箭头表示 Myc 在菠萝蜜 (D. ananassae) 和果蝇 (D.melanogaster) 中位于正 (+) 链上。指向与 Myc 相同方向的宽基因箭头相对于细箭头位于同一链上,而指向 Myc 相反方向的宽基因箭头相对于细箭头位于相反链上。果蝇 (D. ananassae) 中的白色基因箭头表示与果蝇 (D. melanogaster) 中相应基因的直系同源性。 D. ananassae 基因箭头中给出的基因符号表示 D. melanogaster 中的直系同源基因,而基因座标识符特定于 D. ananassae。(B)GEP UCSC Track Data Hub 中的基因模型(Raney 等人,2014 年)。D. ananassae 中 Myc 的编码区显示在用户提供的 Track(黑色)中;CDS 用粗矩形表示,内含子用细线表示,箭头表示转录方向。后续证据轨迹包括 NCBI RefSeq 基因的 BLAT 比对(深蓝色,D. ananassae 的 Ref-Seq 基因比对)、D. melanogaster 蛋白质的 Spaln(紫色,D. melanogaster 的 Ref-Seq 蛋白质比对)、TransDecoder 预测的转录本和编码区(深绿色)、成年雌性、成年雄性和沃尔巴克氏体治愈胚胎的 RNA-Seq(分别为红色、浅蓝色和粉色;D. ananassae 的 Illumina RNA-Seq 读数比对)以及使用 D. ananassae RNA-Seq 由 regtools 预测的剪接点(Graveley 等人,2011;SRP006203、SRP007906;PRJNA257286、PRJNA388952)。显示的剪接点的读取深度 >1000,支持读取为红色。(C)果蝇 Myc-PB 的点图(x 轴)与

果蝇 Pten 直系同源基因模型
(A) 果蝇 (Drosophila melanogaster) 和果蝇 (D. miranda) 中 Pten 基因组邻域的同源性比较。细箭头表示果蝇 (D. melanogaster) (上) 和果蝇 (D. miranda) (下) 中目标基因 Pten 所在的 DNA 链。指向右侧的细箭头表示 Pten 在果蝇 (D. miranda) 中位于正 (+) 链上,指向左侧的细箭头表示 Pten 在果蝇 (D. melanogaster) 中位于负 (-) 链上。指向与 Pten 相同方向的宽基因箭头相对于细箭头位于同一链上,而指向 Pten 反方向的宽基因箭头相对于细箭头位于反链上。果蝇 (D. miranda) 中的白色基因箭头表示与果蝇 (D. melanogaster) 中相应基因直系同源,而黑色基因箭头表示非直系同源。灰色箭头表示在两个基因组邻域中都存在但不是同源的基因(在本例中为 Ror),在 D. miranda 中位于 Pten 的上游,但在 D. melanogaster 中位于 Pten 的下游。D. miranda 基因箭头中给出的基因符号表示 D. melanogaster 中的直系同源基因,而基因座标识符是 D. miranda 特有的。(B)GEP UCSC Track Data Hub 中的基因模型(Raney 等人,2014)。D. miranda 中 Pten 的编码区显示在用户提供的轨道(黑色)中;CDS 用粗矩形表示,内含子用细线表示,箭头表示转录方向。后续证据轨迹包括果蝇 (D. melanogaster) 蛋白质的 Spaln(紫色,果蝇 (D. melanogaster) 的 Ref-Seq 蛋白质比对)、NCBI RefSeq 基因的 BLAT 比对(深蓝色,果蝇 (D. miranda) 的 Ref-Seq 基因比对)、TransDecoder 预测的转录本和编码区(深绿色)、成年雌性和成年雄性的 RNA-Seq(分别为红色和浅蓝色;果蝇 (D. miranda) 的 Illumina RNA-Seq 读段比对)以及使用果蝇 (D. miranda) RNA-Seq (SRP009365) 由 regtools 预测的剪接点。所示的剪接点具有最小读取深度 10,其中 10-49、50-99 和 100-499 支持读取分别以蓝色、绿色和粉色表示。 (C) 果蝇 Pten-PB(x 轴)与果蝇直系同源肽(y 轴)的点图。左侧和底部标明氨基酸编号;顶部和右侧标明 CDS 编号,CDS 也以交替颜色突出显示。点图中的间隙表示序列相似性较低的区域。

人类大脑功能亚群的同源景观...
1 宾夕法尼亚大学佩雷尔曼医学院生物统计学、流行病学和信息学系,宾夕法尼亚州费城 19104,美国;duongtra@usna.edu(DD-T.);jiongc@seas.upenn.edu(JC);sumita.garai@pennmedicine.upenn.edu(SG);fredxu@seas.upenn.edu(FHX);jingxuan.bao@pennmedicine.upenn.edu(JB)2 美国海军学院数学系,马里兰州安纳波利斯 21402,美国 3 普渡大学数学系,印第安纳州西拉斐特 47907,美国; rkaufman@math.purdue.edu 4 宾夕法尼亚大学工程与应用科学学院生物工程系,宾夕法尼亚州,PA 19104,美国 5 乔治梅森大学电气与计算机工程系,弗吉尼亚州费尔法克斯 22030,美国;xwang64@gmu.edu 6 瑞士联邦理工学院洛桑 Neuro-X 研究所,瑞士日内瓦 1015 7 日内瓦大学放射学和医学信息学系,瑞士日内瓦 1211 8 劳伦斯利弗莫尔国家实验室计算工程部,加利福尼亚州利弗莫尔 94550,美国;kaplan7@llnl.gov 9 CENTAI 研究所,意大利都灵 10138; giovanni.petri@nulondon.ac.uk 10 伦敦东北大学网络科学研究所 NPLab,英国伦敦 E1W 1LP 11 IMT 卢卡研究所网络部,意大利卢卡 55100 12 普渡大学综合神经科学研究所,美国印第安纳州西拉斐特 47907;jgonicor@purdue.edu 13 普渡大学工业工程学院,美国印第安纳州西拉斐特 47907 14 普渡大学韦尔登生物医学工程学院,美国印第安纳州西拉斐特 47907 15 耶鲁大学公共卫生学院,美国康涅狄格州新黑文 06520;yize.zhao@yale.edu * 通信地址:li.shen@pennmedicine.upenn.edu † 上述作者对本文的贡献相同。

非同源末端连接是 CRISPR/Cas 的关键……
尽管成簇的规律间隔短回文重复序列 (CRISPR)/CRISPR 相关蛋白 (Cas) 介导的基因编辑已经彻底改变了生物学和植物育种,但大规模的可遗传植物染色体重组仍处于起步阶段。现在可以实现染色体内的重复和倒位,以及染色体之间的易位。随后,可以破坏或新建遗传连锁。此外,染色体上基因的顺序也可以改变。虽然自然染色体重组在减数分裂过程中通过同源重组发生,但 CRISPR/Cas 介导的染色体重排最好通过利用体细胞中的非同源末端连接 (NHEJ) 途径获得。NHEJ 可细分为经典 (cNHEJ) 和替代 NHEJ (aNHEJ) 途径,它们部分地以拮抗方式运作。 cNHEJ 通路不仅可以保护断裂的 DNA 末端免于降解,还可以抑制先前未连接的断裂末端的连接。因此,在没有 cNHEJ 的情况下,可以获得更多的倒位或易位,这可以归因于无限制地使用 aNHEJ 通路进行双链断裂 (DSB) 修复。与倒位或易位相反,短串联重复可以通过 Cas9 切口酶由成对的单链断裂产生。有趣的是,cNHEJ 通路对于这些类型的重复至关重要,而 aNHEJ 则是补丁插入所必需的,补丁插入也可以在 DSB 修复期间形成。由于染色体工程不仅在模式植物拟南芥 (Arabidopsis thaliana) 中实现,而且在作物玉米 (Zea mays) 中也实现,我们预计这项技术将很快改变育种过程。

CRISPR/CAS通过同源重组的第3章基因组编辑
在整个细胞发育中,DNA可能遭受威胁基因组完整性和细胞存活的损害。最有害的病变之一是双链DNA断裂(DSB),因为它可能导致基因组信息的丢失。DSB可能自然发生在细胞代谢期间,也可能是由外部因素触发的(Deriano; Roth,2013)。无论哪种方式,这些损坏都会通过细胞立即修复,主要是通过两种途径:非同源末端连接(NHEJ)或同源指导修复(HDR)。与通过NHEJ进行修复不同,NHEJ仅将裂解的DNA的末端连接起来(请参阅第2章),HDR途径需要存在相同或非常相似的模板,即完整的序列,以准确地修复病变的DNA(Heyer等人,2010年)。提供用于HDR中使用的模板的可能性代表了通过同源重组(HR)途径进行基因编辑的关键元素,该途径可能被利用为几种新的繁殖技术(NBT)之一。

古细菌DNA-Import仪器与细菌共轭机械同源
结合是水平基因转移的主要机制,促进了抗生素耐药性在人类病原体中的传播。它涉及通过称为交配菌毛的细胞外附属物来避免供体和受体细胞之间的连接。在细菌中,结合机制由质粒或转座子编码,通常介导同源移动遗传元件的转移。对古细菌的共轭知之甚少。在这里,我们通过三个共轭pili的冷冻电子显微镜确定原子结构,两种来自高疗法古细菌(Aeropyrum pernix和pyrobaculum calidifontis),另一个由一个由细菌的细菌ti toumefaciial to to to to to to to to to to to to to to toumefacial-to to to to to to to to to to toumefiti。 pili。然而,古细菌共轭机制(称为CED)已被“驯化”,即结合机械的基因编码在染色体上,而不是在移动遗传元素上,并介导细胞DNA的转移。

通过CRISPR介导的同源指导修复的基因标记。
对一些模型植物 - 病原系统的研究已从多年的工具和资源开发中受益。对于绝大多数经济和营养重要的植物而言,情况并非如此,从而产生了农作物改善的瓶颈。木薯细菌疫病(CBB),由xanthomonas axonopodis PV引起。manihotis(XAM)是木薯(Manihot esculenta crantz)种植的所有地区的重要疾病。在这里,我们描述了木薯的开发,可用于可视化体内CBB感染的初始步骤之一。使用CRISPR介导的同源指导修复(HDR),我们在CBB易感性的3'端(S)基因Mesweet10a生成了含有GFP的植物。随后在转录和翻译水平上可视化了转录激活剂(TAL)效应tal20的Mesweet10a-GFP。据我们所知,这是通过木薯中的基因编辑进行HDR的第一个证明。