XiaoMi-AI文件搜索系统
World File Search System对光

液态金属颗粒的尺寸影响液晶弹性体复合材料的驱动性能
对于不依赖环境加热的驱动,LCE 已被合成/加工以响应光、电场或焦耳加热。15 对光的主动响应可能非常快 16,17 并且显然对许多应用有用 18–20 但在远程/无法访问的环境中或在环境光可能影响驱动的情况下可能被禁止。或者,可以使用电场来驱动 LCE。通过添加碳纳米管,LCE 的机电响应性得到改善;然而,相对于未填充的 LCE,刚性内含物会降低驱动应变。21,22 对于厚度大于几百微米的人造肌肉,光和电场驱动都难以扩展。焦耳加热已通过表面加热器和导电填料 23–28 实现,这对于开发使用 LCE 作为软致动器的不受束缚的软机器人很有希望。29,30
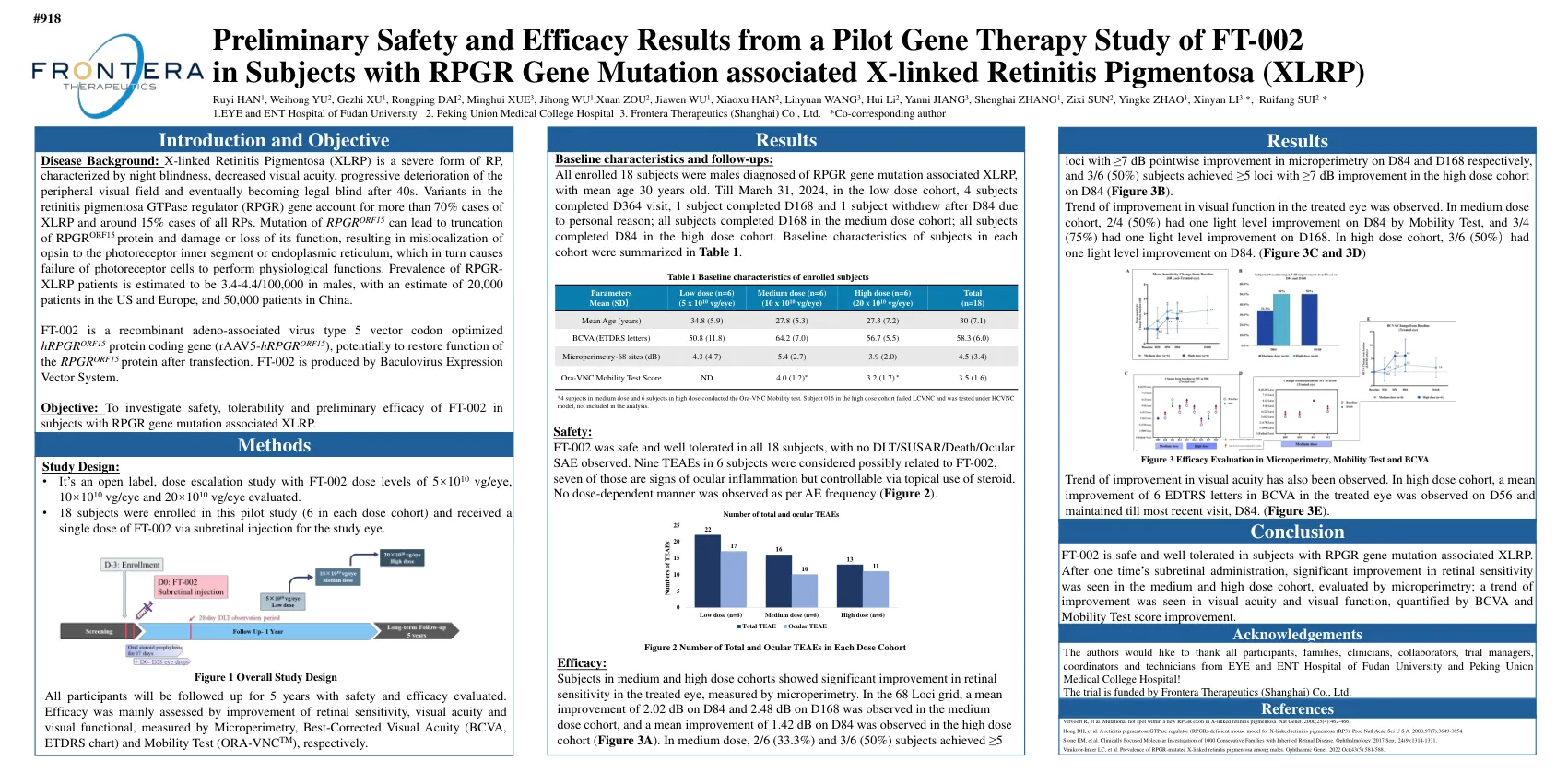
FT-002的试点基因治疗研究的初步安全性和功效引起的RPGR基因突变相关的X连锁性视网膜炎
疾病背景:X连锁性视网膜炎色素(XLRP)是一种严重的RP形式,其特征是夜失明,视力降低,外围视野的进行性恶化,并在40多岁后最终成为法律上的盲人。色素炎的变体GTPase调节剂(RPGR)基因占XLRP的70%以上,所有RPS的病例约为15%。RPGR ORF15的突变会导致RPGR ORF15蛋白质的截断以及其功能的损害或损失,从而导致OPSIN对光感受器内部节段或内质网的错误定位,这反过来会导致光感受器细胞的失败至关重要。RPGR-XLRP患者的患病率估计为男性为3.4-4.4/100,000,在美国和欧洲估计有20,000名患者,在中国有50,000名患者。

水平的眼球震颤带有速度增强的波形,催眠后白血病术中的波形:病例报告
心脏病和糖尿病因眼睛运动异常而被转诊至神经科。在手术后三个月内,他接受了康德疗法,并因甲状腺弥漫性轴突损伤而受到弥散的轴突损伤,并因交通碰撞引起的宫颈椎间盘破裂。最近,由于心脏AR猝死引起的缺氧 - 缺血性脑损伤,该患者表现出异常的眼球运动。神经系统检查表明,他的眼睛处于营养状态,头部固定向左转,四肢瘫痪。学生的大小正常,并且对光反应。角膜和堵嘴反射也是双侧完整的。视频记录术显示,左侧的眼球震颤和小小的下滑,并在患者的角度(从患者的角度来看)扭转组件。去除光不会导致重大变化(图1a,补充视频
光遗传学:用光控制细胞功能
调节膜电位的工具 光遗传学最常见的用途之一是改变可兴奋细胞的膜电位。在神经元中,膜去极化会导致瞬态电信号(脉冲)的激活,这是神经元通讯的基础。相反,膜超极化会导致这些信号的抑制。控制操作这些电流的“开关”使神经科学家能够研究神经元在功能上如何相互关联以及神经元回路如何控制行为。通过外源表达改变神经元膜电位的光激活蛋白,光可以用作开关。一种方法是使用化学修饰的所谓“笼状配体”,这些配体在光刺激下变得活跃并与通过基因引入特定神经元的外源性受体结合。配体也可以通过充当光开关的光敏化合物与受体本身相连。在这两种情况下,都必须将光敏的可溶性或束缚配体注入细胞或组织,使它们对光敏感。或者,可以使用编码光敏蛋白(如视蛋白)的天然基因。这些光敏跨膜蛋白与发色团视网膜共价结合,视网膜吸收光后发生异构化(例如,从反式变为顺式构型),从而激活蛋白质。值得注意的是,视网膜化合物在大多数脊椎动物细胞中含量充足,因此无需注入外源分子。第一个利用视蛋白进行哺乳动物神经元光学控制的遗传编码系统是通过外源表达果蝇的三基因系统建立的。表达这些蛋白质的神经元对光的反应是数秒内的去极化和尖峰波。最近发现,微生物中的视蛋白(将光敏域与同一蛋白质中的离子通道或泵相结合)也可以调节神经元信号,通过在单个易于表达的蛋白质中提供更快的控制,彻底改变了该方法。这些神经元开关中的第一个使用了通道视紫红质-2 (ChR2)。当在神经元中表达并暴露于蓝光时,这种非选择性阳离子通道会立即使神经元去极化
![arxiv:2502.14564v1 [Math.ds] 2025年2月20日](/simg/0\033bab8b12eab1567f556d1dc4884e17af2868bb.webp)
arxiv:2502.14564v1 [Math.ds] 2025年2月20日
BioContightion是一种流动动力学现象,该现象是由比其周围的流体略稠密的自属性微生物的集体运动驱动的。这个过程在各种生物学和工业应用中起着至关重要的作用[1-5]。通气微生物,例如藻类和细菌,在对外部刺激作出反应时会产生密度变化,即一种称为出租车的行为,导致对流稳定性。出租车的关键例子包括照照,重力,陀螺赛,趋化性和趋化性。了解生物对流在环境科学,生物技术和工程学中特别相关,它影响了营养运输,生物反应器效率和微生物生态学。早期研究主要集中于等温条件下的悬浮液。然而,许多微生物,居住在温泉中的良性嗜热剂,在温度变化显着的环境中壮成长[6-8]。在各种类型的出租车中影响微生物运动,光疗(对光的响应)和热疗(对温度梯度的反应)在塑造生物感染模式中起着至关重要的作用[9]。虽然已经针对非孔培养基中的光疗法和引力生物传染进行了大量研究,但充满藻类悬浮液饱和的多孔生物反射仍相对较低。存在多孔矩阵的存在引入了添加复杂性,例如流动性和修饰的构造动力学,使其成为自然生态系统和工业流动系统系统的关键研究领域。但是,当G超过G C时,它们伴有摄影影响的生物配分模式的形成和特征取决于各种环境光条件,包括直接和倾斜的类似的辐射[10-16]。高强度的光可以破坏已建立的模式或抑制其发育[12,13,17]。照明水平的变化有助于这些模式的空间结构和大小的变化。这些改变可以归因于特定机制。光合作用的杂种生物表现出对光强度的方向运动。当强度G保持低于鉴定阈值G C时,它们会表现出正光的阳性,向更明亮的区域迁移。

血浆极性由碱性沉积诱导的Hafnium disulfide $ 1T \ textrm { - } {\ rm hfs} _2 _2 $
我们将基于多体扰动理论和累积膨胀的AB从头算计算与角度分辨光发射光谱(ARPES)相结合,以量化高度掺杂的半导体过渡金属二核基因1 T -HFS中的电子样本相互作用。arpes揭示了传导带底部的准颗粒激发附近的卫星光谱特征的出现,这表明偶联与200 MeV的特征能量的玻体激发偶联。我们对光发射光谱函数的第一个原理计算表明,这些特征可以归因于电子耦合到载体等离子(掺杂诱导的集体电荷密度频率)。我们进一步表明,在表面上减少筛选会增强电子 - 种类的相互作用,并主要负责等离激子极性子的出现。



Malik,Mohammad Ibrahim,Latif,Siddique,Jurdak,Raja和Schuller,BjörnW。(2023)
反映了Giacomo Ciamician革命性的利用阳光以推动光化学转换的革命性愿景,材料科学领域已经显着发展,但由于误解,在吸收最大值时,它在光化学系统中的最高反应性得到了限制。在这里,我们通过光化学作用图的证据进一步探索了这一概念,表明在与吸收峰显着分离的波长下,反应性确实可以是最大的。通过检查吸收性和光化学重新效率之间差异的含义,我们探索了它对光震毒剂中光渗透深度增强的影响,光化学反应的能量需求的减少以及其对体积3D印刷的变换潜力。最终,我们主张对Light促进整个材料的光化学反应的能力的重新欣赏。
