XiaoMi-AI文件搜索系统
World File Search System弥散

补充信息
为了考虑 3d 电子的强相关性并避免局部密度近似中预测的 d 态过度离域,对 Mn 和 Co 分别采用了类 Hubbard 校正 U = 6 eV 和 U = 4 eV(LDA+U 方法)。5 Kampert 等人在计算 {Mn 4 } 时也使用了相同的 U = 6 eV 值。6 对碳、氮和氢使用标准双 zeta 极化 (DZP) 基组,对 Mn、Co 和 O 使用优化的双 zeta (DZ)。计算是自旋极化的,并假设共线自旋。为了确定轨道矩和 SOC 的作用,进行了没有 Hubbard 校正的 LDA+SOC 计算(参考文献 [7] 的场外形式),因为目前的 SIESTA 代码不允许同时包含 SOC 和 Hubbard 校正。我们验证了自旋轨道相互作用的影响在{Mn 4 }中可以忽略不计(对于半填充的3d壳层而言如此),但在{Co 4 }中则不然。在LDA+U计算中,当真实空间网格截止值为400 Ry、费米-狄拉克弥散为100 K时,电子结构和磁性达到了收敛,而在SOC中,截止值为650 Ry,电子温度为1 K。在标准周期边界条件模拟中放宽原子位置,对15个{M 4 }-CNT单元(移位网格)的布里渊区进行1×1×12 k点采样,采用共轭梯度算法。模拟单元沿周期方向延伸36.9354 Å(30个碳原子),而在垂直于管轴的两个方向上,系统的周期复制品之间的真空度超过30 Å。对于 CNT+ {M 4 } 系统,原子上的最大力小于 0.04 eV/Å。开放系统模拟是在非平衡格林函数形式内进行的,使用 TranSIESTA 解决方案方法,8-9 在一个 70 个碳长的单元上进行,该单元由松弛的 {M 4 -CNT} 单元组成,两侧填充有 (5,5)-CNT 片段(总共 20 个碳长)。

559_Palmer_2004_DevelopmentOfAEuropeanMultimodel_Artzeit_pubid11720
热带太平洋 (McPhaden 等人1998),最重要的是,原型耦合海洋-大气模型 (Zebiak 和 Cane 1987) 成功预测了厄尔尼诺现象。反过来,这些发展又得到了非常成功的国际热带海洋全球大气 (TOGA) 计划 (世界气候研究计划 1985) 的推动。季节性预报显然对社会各阶层都有价值,无论是出于个人、商业还是人道主义原因 (例如,Stern 和 Easterling 1999;Thomson 等人2000;Pielke 和 Carbone 2002;Hartmann 等人2002a;Murnane 等人2002)。然而,尽管大气-海洋耦合产生了可预测的信号,但上层大气本质上是混乱的,这意味着预测的天气的日常演变必然对初始条件敏感(Palmer 1993;Shukla 1998)。在实践中,这种敏感性的影响可以通过整合耦合海洋-大气模型的预测的向前时间集合来确定,集合中的各个成员因大气和底层海洋的起始条件的微小扰动而不同。集合的相空间弥散给出了流动潜在可预测性的可量化流动相关测量。但是,如果初始条件的不确定性是季节性预报集合中唯一的扰动,那么由此得出的可预报性测量结果将不可靠;原因是模型方程也是不确定的。更具体地说,尽管气候演变方程在偏微分方程的层面上很容易理解,但它们作为一组有限维常微分方程的表示,在数字计算机上进行积分时,不可避免地会带来不准确性。原则上,这种不准确性可以向上传播,并影响模型所预测的整个尺度范围。目前,没有潜在的理论形式主义可以用来估计模型不确定性的概率分布(见 Palmer 2001);因此,必须寻求一种更务实的方法。其中一种方法依赖于这样一个事实,即全球气候模型是在不同的气候研究所独立开发的。由这种准独立模型组成的集合称为多模型集合。多模型集合能够比单一模型集合产生更可靠的季节性气候风险概率预报,这一点已由季节至年际时间尺度气候变化预测 (PROVOST) 项目资助
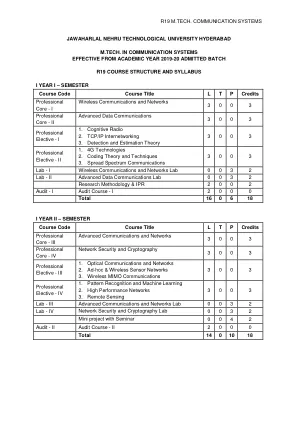
r19 m.tech. 通信系统
单元 I 蜂窝概念系统设计基础:简介、频率重用、信道分配策略、切换策略 - 优先切换、实际切换考虑、干扰和系统容量 - 同信道干扰和系统容量、无线系统的信道规划、相邻信道干扰、减少干扰的功率控制、中继和服务等级、改善蜂窝系统的覆盖范围和容量 - 小区分裂、扇区划分。第二单元移动无线电传播:大规模路径损耗:无线电波传播简介、自由空间传播模型、功率与电场的关系、三种基本传播机制、反射-电介质反射、布儒斯特角、完美导体反射、地面反射(双射线)模型、衍射-菲涅尔区几何、刀刃衍射模型、多重刀刃衍射、散射、室外传播模型-Longley-Ryce 模型、Okumura 模型、Hata 模型、Hata 模型的 PCS 扩展、Walfisch 和 Bertoni 模型、宽带 PCS 微蜂窝模型、室内传播模型-分区损耗(同一楼层)、楼层间分区损耗、对数距离路径损耗模型、爱立信多断点模型、衰减因子模型、信号穿透建筑物、射线追踪和场地特定建模。第三单元移动无线电传播:小规模衰落和多径:小规模多径传播-影响小规模衰落的因素、多普勒频移、多径信道的脉冲响应模型-带宽和接收功率之间的关系、小规模多径测量-直接射频脉冲系统、扩频滑动相关器信道探测、频域信道探测、移动多径信道参数-时间弥散参数、相干带宽、多普勒扩展和相干时间、小规模衰落的类型-由于多径时间延迟扩展而导致的衰落效应、平坦衰落、频率选择性衰落、由于多普勒扩展而导致的衰落效应-快速衰落、慢速衰落、多径衰落信道的统计模型-Clarke 的平坦衰落模型、Clarke 模型中由于多普勒扩展而导致的频谱形状、Clarke 和 Gans 衰落模型的模拟、电平交叉和衰落统计、双射线瑞利衰落模型。
SIFT2:利用流线纤维束成像技术对大脑白质连接进行密集定量评估
目标受众:对使用扩散 MRI 流线纤维束成像定量评估大脑白质连接感兴趣的研究人员。目的:由于流线重建过程的非定量性质 [1],使用扩散 MRI 定量评估大脑白质连接非常困难。针对该问题提出的解决方案包括启发式校正已知的重建偏差 [2,3](可能无法补偿所有重建误差)或评估连接路径上某些扩散模型参数 [4,5,6](依赖于该参数的量化和可解释性)。最近,提出了球面反卷积信息纤维束成像滤波 (SIFT) 方法 [7],通过选择性去除流线,将重建的流线密度与通过扩散信号球面反卷积估计的单个纤维群体积 [8] 进行匹配;完成此过程后,连接两个区域的流线计数变为连接这些区域的白质通路横截面积的估计值(最高可达全局缩放因子)。之前已证明,如果首先应用 SIFT 方法 [9],大脑连接的定量测量与从人脑解剖估计的特性会更加密切相关。这种方法的缺点是,即使生成了许多流线(计算成本高昂),完成过滤后,流线密度可能非常低(这对于定量分析来说是不可取的 [10,11])。在这里,我们提出了一种替代解决方案,称为 SIFT2:此方法不是去除流线,而是为每条流线得出合适的加权因子,以使总流线重建与测量的扩散信号相匹配。方法:与原始 SIFT 方法一样,我们执行纤维方向分布 (FOD) 分割,将流线分配给它们穿过的 FOD 叶,并得出一个处理掩模,以减少非白质体素对模型的贡献。我们将离散 FOD 叶 L 的积分表示为 FOD L ,将归因于该叶的流线密度表示为 TD L ,将处理掩模 [7] 在该叶所占体素中的值表示为 PM L ;从这些中我们得出比例系数 μ [7](等式 1)。每条流线 S 都有一个关联的加权系数 FS 。FOD 叶 L 中的流线密度定义为(等式 2),其中 | SL | 是流线 S 穿过归因于 FOD 叶 L 的体素的长度。目标是找到一组加权系数 FS ,以最小化成本函数 f(等式 3),其中 λ 是用户可选择的正则化乘数,它将流线加权系数约束为与穿过相同 FOD 叶的其他流线相似(等式 4)。使用迭代线搜索算法可以找到解决方案:每个加权系数都经过独立优化,同时考虑一组相关项,这些相关项表示在对每个系数进行独立牛顿更新的情况下所有 L 的 TD L 的估计变化(等式 5)。数据采集和预处理:图像数据是从健康男性志愿者的 3T Siemens Tim Trio 系统(德国埃尔朗根)上采集的。DWI 协议如下:60 个弥散敏化方向,b =3,000s.mm -2,7 b =0 体积,60 个切片,2.5mm 各向同性体素。使用 MPRAGE 序列(TE/TI/TR = 2.6/900/1900ms,9° 翻转,0.9mm 各向同性体素)获取解剖 T1 加权图像。对弥散图像进行了校正以适应受试者运动 [12]、磁化率引起的扭曲 [13] 和 B 1 偏置场 [14]。使用约束球面反卷积 (CSD) [15] 估计纤维取向分布。使用 iFOD2 概率流线算法 [16] 生成了 1000 万条流线的纤维束图,该算法结合了解剖约束纤维束成像框架 [17] ,随机分布在整个白质中。结果:将 SIFT2 与执行 SIFT“收敛”(移除尽可能多的流线以实现与数据的最佳拟合 [7] )进行了比较。对于 SIFT2,我们使用了 λ = 0.001,这是基于近似 L 曲线分析选择的。SIFT 和 SIFT2 方法都以这样一种方式操纵重建,使得流线密度与通过 CSD 得出的体积估计值高度一致(图 1)。然而,SIFT2 实现了比 SIFT 更优秀的模型拟合,同时保留了初始重建中的所有流线(而 SIFT 必须去除大约 96% 的流线)。根据近似 L 曲线分析选择。SIFT 和 SIFT2 方法都以流线密度与通过 CSD 得出的体积估计值高度一致的方式操纵重建(图 1)。然而,SIFT2 实现了比 SIFT 更好的模型拟合,同时保留了初始重建中的所有流线(而 SIFT 必须删除大约 96% 的所有流线)。根据近似 L 曲线分析选择。SIFT 和 SIFT2 方法都以流线密度与通过 CSD 得出的体积估计值高度一致的方式操纵重建(图 1)。然而,SIFT2 实现了比 SIFT 更好的模型拟合,同时保留了初始重建中的所有流线(而 SIFT 必须删除大约 96% 的所有流线)。

SGLT-2 抑制剂作为二甲双胍的附加治疗对 2 型糖尿病患者 P 波指数和心房电机械的影响
缩写:LA,左心房;LAVI,左心房容积指数;IEMD,心房间电机械延迟;LEMD,左侧心房内电机械延迟;LVEF,左心室射血分数;PA,P 至 A' 间期;PTFV1,V1 中的 P 终末力;PWD,P 波弥散;REMD,右侧心房内电机械延迟。 *I 组 vs. III 组;p = .035。**I 组 vs. III 组 p = .002。***I 组 vs. II 组;p = .042,I 组 vs. III 组;p < .001,II 组 vs. III 组;p = .033。****I 组 vs. II 组;p = .011,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .005。*****I 组 vs. II 组; p = .035,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .005。+ I 组 vs. II 组;p = .042,I 组 vs. III 组;p < .001,II 组 vs. III 组;p = .021。++ I 组 vs. II 组;p = .012,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .004。+++ I 组 vs. II 组;p = .002,I 组 vs. III 组;p < .0001,II 组 vs. III 组;p < .001。¥ I 组 vs. III 组;p < .0001,II 组 vs. III 组;p = .038。¥¥ I 组 vs. II 组;p = .026,I 组 vs. III 组; p < .0001,II 组与 III 组;p = .023。¥¥¥ I 组与 II 组;p = .016,I 组与 III 组;p < .0001,II 组与 III 组;p = .036。§ I 组与 II 组;p = .042,I 组与 III 组;p < .001,II 组与 III 组;p = .039。§§ I 组与 II 组;p = .044,I 组与 III 组;p = .003,II 组与 III 组;p = .040。

大脑通讯 AIN 通讯
帕金森病的早期和准确鉴别诊断仍然是临床医生面临的重大挑战。近年来,许多研究利用磁共振成像数据结合机器学习和统计分类器成功区分了不同形式的帕金森病。然而,为了尽量减少偏差和伪影驱动的分类,仍存在一些问题和方法问题。在本研究中,我们比较了不同的特征选择方法和不同的磁共振成像模式,并匹配良好的患者组,并严格控制与患者运动相关的数据质量问题。我们的样本来自 69 名健康对照者,以及特发性帕金森病 (n = 35)、进行性核上性麻痹理查森综合征 (n = 52) 和皮层基底节综合征 (n = 36) 患者。参与者接受了标准化 T1 加权和弥散加权磁共振成像。严格的数据质量控制和组匹配将对照组和患者组的数量分别减少到43、32、33 和 26。我们比较了两种不同的特征选择和降维方法:全脑主成分分析和基于解剖感兴趣区域的方法。在这两种情况下,支持向量机都用于构建健康对照组和患者的成对分类的统计模型。使用留二交叉验证方法以及使用不同受试者集的独立验证来估计每个模型的准确度。我们的交叉验证结果表明,与基于感兴趣区域的方法相比,使用主成分分析进行特征提取可提供更高的分类准确度。然而,当使用独立样本进行验证时,两种特征提取方法之间的差异显著缩小,这表明主成分分析方法可能更容易受到交叉验证过度拟合的影响。 T1 加权和扩散磁共振成像数据均可用于成功区分受试者组,在交叉验证分析的所有成对比较中,两种方式均不优于另一种方式。但是,当使用独立验证队列时,从扩散磁共振成像数据获得的特征可显著提高分类准确率。总体而言,我们的结果支持使用统计分类方法对帕金森病进行鉴别诊断。但是,分类准确率可能会受到组大小、年龄、性别和运动伪影的影响。通过适当的控制和样本外交叉验证,包括基于磁共振成像的分类器在内的诊断生物标志物评估可能是临床评估的重要辅助手段。
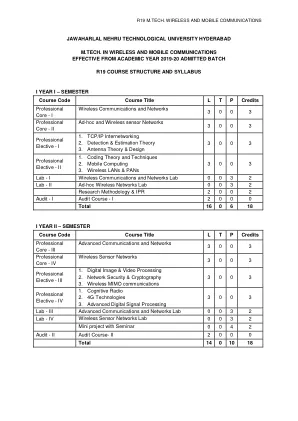
r19 m.tech. 无线和移动通信
1. 理解蜂窝通信概念 2. 研究移动无线电传播 3. 研究无线网络不同类型的 MAC 协议 UNIT -I 蜂窝概念-系统设计基础:简介、频率重用、信道分配策略、切换策略 - 优先切换、实际切换考虑、干扰和系统容量 - 同信道干扰和系统容量、无线系统的信道规划、相邻信道干扰、减少干扰的功率控制、中继和服务等级、提高蜂窝系统的覆盖范围和容量 - 小区分裂、扇区划分。第二单元移动无线电传播:大规模路径损耗:无线电波传播简介、自由空间传播模型、功率与电场的关系、三种基本传播机制、反射-电介质反射、布儒斯特角、完美导体反射、地面反射(双射线)模型、衍射-菲涅尔区几何、刀刃衍射模型、多重刀刃衍射、散射、室外传播模型-Longley-Ryce 模型、Okumura 模型、Hata 模型、Hata 模型的 PCS 扩展、Walfisch 和 Bertoni 模型、宽带 PCS 微蜂窝模型、室内传播模型-分区损耗(同一楼层)、楼层间分区损耗、对数距离路径损耗模型、爱立信多断点模型、衰减因子模型、信号穿透建筑物、射线追踪和场地特定建模。第三单元移动无线电传播:小规模衰落和多径:小规模多径传播-影响小规模衰落的因素、多普勒频移、多径信道的脉冲响应模型-带宽和接收功率之间的关系、小规模多径测量-直接射频脉冲系统、扩频滑动相关器信道探测、频域信道探测、移动多径信道参数-时间弥散参数、相干带宽、多普勒扩展和相干时间、小规模衰落的类型-由于多径时间延迟扩展而导致的衰落效应、平坦衰落、频率选择性衰落、由于多普勒扩展而导致的衰落效应-快速衰落、慢速衰落、多径衰落信道的统计模型-Clarke 的平坦衰落模型、Clarke 模型中由于多普勒扩展而导致的频谱形状、Clarke 和 Gans 衰落模型的模拟、电平交叉和衰落统计、双射线瑞利衰落模型。第四单元均衡和分集:介绍、均衡基础知识、训练通用自适应均衡器、通信接收器中的均衡器、线性均衡器、非线性均衡器

多模态磁共振在缺血性脑卒中评估中的运用进展
[1] Murray CJL, Aravkin AY, Zheng P, et al.Global burden of 87 risk factors in 204 countries and territories, 1990 – 2019: a systematic analysis for the Global Burden of Disease Study 2019[J].Lancet, 2020, 396 (10258): 1223-1249.[2] 王增武 , 马志毅 , 薛素芳 , 等 .基层冠心病与缺血性脑卒中共患管理 专家共识 2022[J].中国心血管病研究 , 2022, 20(9): 772-793.[3] 王拥军 , 李子孝 , 谷鸿秋 , 等 .中国卒中报告 2020 (中文版) (1)[J].中 国卒中杂志 , 2022, 17(5): 433-447.[4] Langhorne P, Bernhardt J, Kwakkel G. Stroke rehabilitation[J].Lancet, 2011, 377(9778): 1693-1702.[5] Xing Y, Bai Y.A Review of Exercise-Induced Neuroplasticity in Ischemic Stroke: Pathology and Mechanisms[J].Mol Neurobiol, 2020, 57 (10): 4218-4231.[6] Guggisberg AG, Koch PJ, Hummel FC, et al.Brain networks and their relevance for stroke rehabilitation[J].Clin Neurophysiol, 2019, 130(7): 1098-1124.[7] Lutsep HL, Albers GW, Decrespigny A, et al.Clinical utility of diffusion-weighted magnetic resonance imaging in the assessment of ischemic stroke[J].Ann Neurol, 1997, 41(5): 574-580.[8] 于帆 , Arman Sha, 张苗 , 等 .人工智能在急性缺血性脑卒中影像的研 究进展 [J].中华老年心脑血管病杂志 , 2023, 25(3): 334-336.[9] 李华 , 郭春锋 , 高连荣 .FLAIR 及 DWI 序列在诊断脑血管周围间隙 中的价值 [J].医学影像学杂志 , 2015, 25(8): 1341-1343.[10] Scheldeman L, Wouters A, Dupont P, et al.Stroke, 2022, 53(5): 1665-1673.[11] Thomalla G, Simonsen CZ, Boutitie F, et al.MRI-Guided Thrombolysis for Stroke with Unknown Time of Onset[J].[15] 蔡建新 , 彭如臣 .扩散加权成像和流体且反转的恢复定量定量,以预测不明发作的缺血性中风中的易流性恢复不匹配的恢复不匹配状态[J]。《新英格兰医学杂志》,2018,379(7):611-622。[12] Legrand L,Turc G,Edilali M等。根据Flair血管高压不匹配而受益于血栓切除术后血运重建[J]。Eur Radiol,2019,29(10):5567-5576。[13] Xie Y,Oppenheim C,Guillemin F等。预处理病变量会影响临床结果和血栓切除术的功效[J]。Ann Neurol,2018,83(1):178-185。 [14] Raoult H,Lassalle MV,Parat B等。 基于DWI的算法可预测急性中风血栓切除术治疗的患者的残疾[J]。 Am J Neuroradiol,2020,41(2):274-279。 弥散张量磁共振成像方法概述[J]。 医学影像学杂,2007,17(10):1119-1122。 [16] Qiu A,Mori S,Miller MI。 扩散张量成像,用于理解早期生命中大脑发育[J]。 Ann Rev Psychol,2015,66:853-876。 [17] Corroenne R,Arthuis C,Kasprian G等。 胎儿大脑的扩散张量成像:有前途技术的原理,潜力和局限性[J]。 超声产科妇科,2022,60(4):470-476。 [18] Andica C,Kamagata K,Hatano T等。 源自扩散成像的退化性脑疾病的生物标志物[J]。 J Magn Reson Imaging,2020,52(6):1620-1636。 [19] Groisser BN,哥伦WA,Singhal AB等。 NeuroRehabil神经修复,2014,28(8):751-760。Ann Neurol,2018,83(1):178-185。[14] Raoult H,Lassalle MV,Parat B等。基于DWI的算法可预测急性中风血栓切除术治疗的患者的残疾[J]。Am J Neuroradiol,2020,41(2):274-279。弥散张量磁共振成像方法概述[J]。医学影像学杂,2007,17(10):1119-1122。[16] Qiu A,Mori S,Miller MI。扩散张量成像,用于理解早期生命中大脑发育[J]。Ann Rev Psychol,2015,66:853-876。 [17] Corroenne R,Arthuis C,Kasprian G等。 胎儿大脑的扩散张量成像:有前途技术的原理,潜力和局限性[J]。 超声产科妇科,2022,60(4):470-476。 [18] Andica C,Kamagata K,Hatano T等。 源自扩散成像的退化性脑疾病的生物标志物[J]。 J Magn Reson Imaging,2020,52(6):1620-1636。 [19] Groisser BN,哥伦WA,Singhal AB等。 NeuroRehabil神经修复,2014,28(8):751-760。Ann Rev Psychol,2015,66:853-876。[17] Corroenne R,Arthuis C,Kasprian G等。胎儿大脑的扩散张量成像:有前途技术的原理,潜力和局限性[J]。超声产科妇科,2022,60(4):470-476。[18] Andica C,Kamagata K,Hatano T等。源自扩散成像的退化性脑疾病的生物标志物[J]。J Magn Reson Imaging,2020,52(6):1620-1636。[19] Groisser BN,哥伦WA,Singhal AB等。NeuroRehabil神经修复,2014,28(8):751-760。皮质脊髓扩散异常[J]。[20] Kumar P,Kathuria P,Nair P等。使用扩散张量成像的亚急性缺血性卒中后上肢运动恢复的预测:系统评价和荟萃分析[J]。J Stroke,2016,18(1):50-59。[21] Soulard J,Huber C,Baillieul S等。运动道完整性预测步行恢复:亚急性中风中的扩散MRI研究[J]。神经病学,

婴儿颤抖综合征和自闭症
NLM 提供对科学文献的访问,但并不意味着 NLM 或美国国立卫生研究院认可其内容。详细了解 PMC 免责声明和版权声明。2021 年 2 月 1 日发表在 PMC 上的一项研究发现,自闭症谱系障碍 (ASD) 出现在幼儿时期,当时婴儿从正常的行为特征过渡到幼儿期表现出 ASD 特征。前瞻性脑成像研究通过揭示 ASD 的神经生物学和发育过程,显示出在症状前检测和为早期干预提供信息方面的巨大希望。本文回顾了从出生到幼儿期 ASD 大脑发育的神经影像学研究,将这些发现与候选神经生物学机制联系起来,并讨论了对未来研究和临床实践的影响。在美国,ASD 的患病率为 1/59,其特点是症状特征各异,社交沟通障碍和限制性重复行为的严重程度各不相同。尽管人们对了解自闭症的神经生物学非常感兴趣,但大多数研究都是横断面研究和诊断后研究,涉及的年龄范围很广。最近的前瞻性研究跟踪了高风险兄弟姐妹从婴儿期到幼儿期的情况,发现自闭症的诊断症状在生命的第一年和第二年的后半段出现。运动技能、对面部和社交场景的关注、对名字的反应、视觉接收和语言技能的差异在生命第二年的早期也很明显。这些行为发生在出生后大脑发育的高度动态时期,其特点是大脑结构和功能发生重大变化。自闭症谱系障碍 (ASD) 患者的大脑发育已得到广泛研究,研究使用了 MRI 等神经成像技术。研究表明,非典型大脑表型在婴儿期出现,通常在两岁左右症状巩固之前。研究表明,后来患上自闭症的婴儿在 12 至 24 个月之间表现出更快的总脑容量增长速度,与非自闭症同龄人相比,这些个体的脑容量有所增加。最近的研究还将生命第二年期间大脑总体积的变化率与 ASD 相关的社交缺陷的严重程度联系起来。此外,研究表明,大脑过度生长不是出生时存在的,而是在生命第一年的后期出现的。这些发现对临床实践具有重要意义,并强调需要进一步研究以确定个人特定的发育问题领域,利用神经学特征分析病因异质性,将遗传变异纳入神经影像学研究,绘制大脑发育和行为表型的共现图,并将体内 MRI 与基础科学相结合,揭示 ASD 病理生理学的机制见解。研究发现,6 至 12 个月大的婴儿的大脑发育显著增长,后来患上了自闭症谱系障碍 (ASD),并在生命第二年出现大脑过度生长。这一发现支持了皮质过度扩张导致 ASD 大脑过度生长的假设。此外,使用机器学习方法通过 6 个月和 12 个月的 MRI 测量值进行诊断分类。研究还发现皮质表面积和厚度的差异检查,ASD 婴儿和幼儿与对照组之间没有发现差异。一项研究在某些情况下观察到局部皮质区域的厚度增加,这可能是由于年龄范围或使用的图像分析管道造成的。在青少年和成年人中,观察到皮质厚度差异,但影响的方向不同。混合纵向设计发现,对于患有 ASD 的个体,儿童时期的皮质厚度较大,随后在中期轨迹交叉,成年早期局部皮质厚度减少。研究表明,皮质厚度的异常模式在 3 岁后出现,此后遵循动态发展模式。还检查了皮质脑回模式,一项研究发现 3 岁时患有自闭症的男孩的梭状回脑回减少,并且脑回纵向增加。在患有自闭症谱系障碍 (ASD) 的个体中,在学龄前,颞叶、额叶和顶叶等区域的脑回增加,而正常发育的对照组局部脑回保持稳定或减少。这与之前关于患有自闭症的大龄儿童和成人大脑发育增加的发现一致。需要进一步研究来揭示患有自闭症的幼儿和婴儿大脑结构的发育模式。杏仁核是大脑的核心社交区域,引起了人们对自闭症病理生理学的极大兴趣,但很少有研究探索其在儿童早期的发展。研究表明,学龄前杏仁核增大与较差的社交和沟通结果相关,在患有 ASD 的女孩身上观察到了显著的影响。纵向调查揭示了患有 ASD 的幼儿的杏仁核大小、行为和遗传风险因素之间的复杂关系。作者比较了正常发育儿童和发育迟缓儿童的小脑体积,但没有发现行为和小脑体积之间的关联。然而,一项针对患有 ASD 的幼儿的研究报告称,小脑内的白质体积较大,灰质增加,尤其是在女性中。其他研究表明,病例组和对照组的小脑体积没有差异,而一些研究表明,与正常发育个体相比,自闭症儿童和成人的胼胝体可能较小。一项对具有自闭症家族风险的婴儿的纵向研究发现,他们的胼胝体面积在出生后第一年增加,但到 2 岁时就恢复正常。此外,在这些婴儿中还观察到轴外液量的增加,这种增加在患上自闭症之前一直持续到 24 个月。研究发现,6 个月时的轴外液量与自闭症谱系障碍 (ASD) 严重程度有关。在更大的婴儿群体中,与对照组相比,患有自闭症的婴儿轴外液量增加了 18%。该研究还报告称,自闭症症状最严重的儿童轴外液量增加了 25%。Shen 和同事发现,无论孩子是否有家族风险,轴外液的增加都会持续到 3 岁。他们还将体液增加与自闭症儿童的睡眠问题和非语言能力下降联系起来。使用扩散 MRI 的研究调查了 ASD 中的白质连接性和完整性。虽然很少有研究关注学龄前时期,但早期研究结果表明大脑某些区域的分数各向异性 (FA) 较高,表明白质特性更成熟。尽管在很宽的年龄范围内都出现了下降,但研究发现患有自闭症谱系障碍 (ASD) 的幼儿和儿童的分数各向异性 (FA) 较低。两项纵向研究揭示了 ASD 中白质发育的动态发展性质。一项研究跟踪了 6 至 24 个月大有患 ASD 风险的婴儿,发现那些后来患上 ASD 的婴儿最初表现出 FA 增加,随后成熟速度变慢。另一项研究报告了与年龄相关的 FA 异常变化,FA 在较小年龄时较大,后来变化速度变慢。这些发现表明 ASD 的特点是生命第一年 FA 增加,随后成熟速度变慢,最终可能导致年龄较大的儿童和成年人的 FA 值降低。最近的研究还探索了白质发育作为网络或连接组的情况。一项研究发现,与对照组相比,患有 ASD 的幼儿局部和整体效率降低,尤其是在感觉处理区域。另一项研究表明,在后来患上 ASD 的婴儿中,早在 6 个月大时,白质网络效率就存在缺陷。此外,研究将白质发育与幼儿的 ASD 相关行为联系起来,包括限制性和重复性行为以及对感觉刺激的反应。语言分数的个体差异与白质发育的差异有关。对有自闭症谱系障碍 (ASD) 家族风险的婴儿的研究发现,大脑结构的改变可能导致 ASD 的行为紊乱。功能性磁共振成像研究揭示了神经活动对听觉刺激的反应存在差异,包括大脑半球之间的同步性降低和语言网络的异常侧化。与对照组相比,患有自闭症的幼儿表现出较弱的半球间同步性,双侧颞叶和额叶区域的激活度降低。该研究还发现大脑与行为之间的关系呈负相关,表明自闭症患者的语言区域功能特化异常。研究表明,婴儿在患上自闭症谱系障碍 (ASD) 后,某些区域(如扣带回和岛叶)的大脑活动可能会发生变化。然而,还需要更多的研究来证实这些模式是否是自闭症所特有的。研究还发现,患有自闭症的小男孩的杏仁核与其他参与社交沟通和重复行为的大脑区域之间的联系减弱。一项针对有患自闭症风险的婴儿的研究发现,不同大脑网络的功能连接与后来的限制性和重复性行为的发展之间存在关联。然而,随着孩子长大,这种关联的方向发生了变化。研究还表明,静息状态连接可用于预测 6 个月大婴儿的诊断结果。早在 6 个月大时,患上 ASD 的婴儿就表现出异常的白质发育和脑脊液量增加,这与运动延迟和非典型视觉定向相吻合。大脑变化先于定义 ASD 特征的出现,并与生命第一年的行为变化有关。这些发现表明,大脑表型保持稳定,而 ASD 症状在生命第二年巩固。跨多个范式的研究(包括每个表型的纵向研究)支持此处提出的发现(图)。双条表示轨迹的未知或记录不全的起点和/或终点。顶部面板中的虚线表示典型的大脑发育,上下偏差表示相对于对照组的大脑表型增加或减少。例如,与对照组相比,ASD 中的分数各向异性在 6 个月时增加,在 12 个月时保持不变,从 24 个月到 36 个月时降低。重复行为和社交缺陷持续超过 36 个月,没有被引用,因为这些是自闭症患者的诊断特征。第一年的表面积过度扩张先于第二年的大脑过度生长34。同时,对名字的反应改变从 9 个月开始,并持续到 24 个月21,与对照组相比,注意力轨迹不同19,自闭症症状的出现9,11–14。这些发现有助于形成一条发展时间表,其中与自闭症和风险相关的大脑和行为表型在前驱期出现,大致在两岁生日之前,此后诊断症状得到巩固。在灰质发育和皮质表面积扩大的推动下,头两年大脑快速生长27。然而,在 ASD 中,这种出生后的轨迹被打乱了。行为和神经影像学研究表明,皮质表面积的过度扩张与 6 至 12 个月前观察到的运动、感觉和视觉缺陷的前驱期同时发生,随后在第二年出现大脑过度生长和自闭症社交缺陷2。这凸显了控制表面积扩张的机制在 ASD 病理生理学中的核心作用。扩张被认为是由神经祖细胞增殖、分化和迁移113–116 控制的,oRG 细胞群扩张与大脑大小直接相关113。神经祖细胞增殖和神经发生在 ASD 发展中的潜在作用得到了临床前、遗传、尸检数据118 和最近研究的支持,这些研究表明来自 ASD 患者的细胞存在过度增殖。此处给出文章文本 大脑生长加快,特别是某些区域(例如视觉皮层)的大脑生长加快,可能是自闭症综合征的标志,包括 16p11 缺失、PTEN 和 Chd8 突变。研究表明,神经元增殖增加会导致神经连接发生变化,进而影响电路功能和行为。对小鼠的研究还发现,上层锥体神经元的过度繁殖会破坏正常的大脑发育,导致突触连接改变和类似自闭症的行为。此外,研究报告称,患有大头畸形的 ASD 患者的突触形成和神经元兴奋性发生了改变,抑制性神经元和突触也增加了。这些发现表明,大脑生长异常和神经回路中断可能是某些自闭症谱系障碍 (ASD) 亚型的潜在因素。此外,在 ASD 小鼠模型中观察到了经验依赖性可塑性和突触修剪机制的中断,这凸显了平衡兴奋性和抑制性突触在调节神经元之间竞争方面的重要性。研究还表明,6 至 12 个月的皮质过度扩张可能导致视觉定向行为缺陷,最终导致电路修剪效率低下和 ASD 特征的出现。此处给出文章文本自闭症谱系障碍 (ASD) 中存在轴外液体量表明存在超出当前理解的其他致病机制。最近的研究强调了脑脊液 (CSF) 在大脑发育和功能中的作用,研究结果表明脑脊液循环中断和代谢物积累会影响大脑功能。在表面积过度扩张之前增加的脑脊液量表明脑脊液在 ASD 的病理生理学中起调节作用。胼胝体形态和白质微结构的改变暗示着髓鞘形成、轴突口径和连接性控制过程。髓鞘形成基因富集的小鼠模型已发现少突胶质细胞功能发生显著改变,导致髓鞘厚度减少和连接效率低下。这些发现支持了这样一种观点,即 ASD 源于多种产前和产后致病机制,包括神经增殖、迁移、突触发生、修剪、髓鞘形成以及轴突发育和连接。尽早发现婴儿期的 ASD 对开发更有效的治疗方法至关重要。这一过程的复杂性反映在 ASD 患者身上观察到的多种症状和临床结果上。最近的研究使用机器学习技术分析婴儿 MRI 扫描,并以高精度预测 24 个月时的 ASD 诊断。特别是两项研究表明,生命第一年收集的 MRI 扫描可用于准确识别将继续发展为 ASD 的婴儿。一项研究开发了一种深度学习算法,该算法正确预测了 106 名高危婴儿的诊断结果,灵敏度为 88%,特异性为 95%,阳性预测值为 81%。这种方法优于生命头两年的行为测量,并有可能在大脑发育的关键时期实现早期干预。另一项研究使用支持向量回归机在 59 名高危婴儿样本中预测 ASD 诊断,灵敏度为 82%,特异性为 100%,阳性预测值为 100%。这些发现为使用 MRI 和机器学习技术进行更大规模的症状前诊断分类研究铺平了道路。在儿科神经影像学中使用数据驱动方法有望绕过事先进行特征选择的需要,从而实现更准确和更通用的模型。研究表明,深度学习 (DL) 方法可以实现更高的抽象和复杂程度,从而检测数据中的细微模式。然而,在经验丰富的专业人员(如人工智能科学家、统计学家或工程师)的监督下使用这些方法至关重要,他们经常将机器学习算法应用于高维数据集。对疾病的临床了解对于解释这些模型产生的复杂结果也至关重要。进行基于神经影像的预测研究的最佳实践包括确保足够的样本量和普遍性、解释和方法透明度。未来使用大型、公开可用的数据集的工作将有助于解决与样本量和类别不平衡相关的问题。解决这些问题需要开发新方法,例如合成过采样策略。了解从 MR 图像中得出的哪些特征有助于分类也至关重要。虽然目前的方法可以解释深度学习模型,但需要进一步研究来应对这一挑战。最终,在出版物中报告和共享机器学习算法的透明度对于共享知识和为该领域的最佳实践制定标准是必要的。该研究采用了机器学习算法,报告样本量、交叉验证、训练、测试程序、解决类别不平衡、调整参数和优化步骤。应包括解释结果的详细信息,包括识别算法学习的信息和临床相关的性能指标(特异性、敏感性、阳性预测值)。必须提供用于验证和复制的用于构建算法和进行分析的代码。大规模的症状前个性化预测对于塑造临床实践具有重大意义,必须仔细考虑伦理影响。神经科学中从群体层面的相关性到个体层面的预测的转变对于改善生活至关重要,首先是通过将模型应用于新的独立数据集来复制开创性的研究。心理放射学的发展已显示出希望,旨在实现精神疾病的个性化预测。将经过验证的算法整合到临床实践中符合精准医疗框架,为个体分配个性化治疗计划。早期诊断和干预至关重要;虽然存在针对 ASD 的循证行为干预,但预防性干预仍未经证实。神经影像学可以用作基于生物学的筛查工具,指导未来的研究。考虑到 ASD 和神经发育障碍的表型变异性,下一步的主要工作是开发方法来预测个性化关注领域。超过四分之一的有家族性 ASD 风险的婴儿在头几年会出现亚阈值异常行为,使他们成为有针对性干预的候选人。机器学习方法已经证明了使用新生儿扩散 MRI 对幼儿期认知结果的个性化预测。未来的工作应该将类似的方法应用于有 ASD 风险的婴儿。解析神经发育特征中的异质性是一种有前途的方法,可以了解 ASD 等复杂神经精神疾病的症状多样性。 NIMH 的 RDoC 项目专注于根据神经特征识别亚组,以揭示病因和治疗方面的见解。实施聚类算法可以帮助识别疾病的不同轨迹,可能反映不同的病因。虽然遗传研究已经确定了一些 ASD 病例中的新生突变,但常见的多基因变异被认为是大多数病例的原因。可遗传背景遗传变异、多基因风险之间的关系婴儿期和幼儿期大脑发育的特定个体差异以及原因仍然未知。最近对综合征型 ASD 的研究显示了背景遗传变异对幼儿行为发育的预测能力。未来的研究应将其扩展到特发性 ASD,使用神经影像学揭示早期行为表现的见解。患有 ASD 的婴儿表现出各种大脑表型,包括过度生长、液体量增加和白质发育异常,但没有一种足以预测诊断或确定因果机制。为了更好地理解这些表型及其与行为的关系,绘制从婴儿期到诊断期间大脑和行为表型的共同发展过程应该是一个主要的科学目标。先前的研究主要集中于对大脑发育的早期阶段进行建模,但需要更多地关注可能对自闭症谱系障碍 (ASD)185 至关重要的后期阶段。未来的研究应扩展到患有 ASD 和表现出大脑过度生长表型 119–121 的个体之外,以更深入地了解该疾病的根本原因。脑成像数据可以区分有患 ASD 风险的婴儿和正常发育的儿童,甚至在出现任何明显的行为问题之前。许多研究得出了几个关键发现,包括患有 ASD 的个体的脑容量增加、轴外液体量、白质发育改变和连接模式异常。这表明各种神经生物学因素都会影响儿童早期的大脑和行为发育。最近的进展促成了个性化预测模型的开发,用于识别患 ASD 风险较高的婴儿,强调需要有效的症状前干预措施。未来的研究应集中于研究病因异质性,并通过结合神经影像学、行为和基础科学研究的多学科方法将大脑和行为发育与潜在的遗传机制联系起来。该领域在描述婴儿期和幼儿期与自闭症相关的大脑表型方面取得了重大进展,包括大脑过度生长、脑脊液量增加、白质发育改变以及结构和功能连接模式异常。使用神经影像数据预测诊断和维度结果对推进临床实践大有裨益。未来的工作应侧重于解析自闭症的异质性、将遗传变异与脑影像数据联系起来、绘制发育大脑和行为表型的共现图表以及将神经影像研究与基础科学研究相结合。近年来,自闭症早期大脑和行为发育的研究取得了重大进展。研究揭示了从出生到学龄前自闭症症状的出现,神经成像技术揭示了大脑发育的不同模式。这些发现表明自闭症可能与早期大脑结构和功能异常有关。2017 年发表的一项研究提出了一个概念框架,用于理解自闭症早期大脑和行为发育。另一项研究发现,年仅 12 个月大的婴儿表现出重复性行为,这些行为后来成为自闭症谱系障碍 (ASD) 的特征。自闭症遗传学研究也取得了进展,一些研究表明兄弟姐妹中自闭症复发风险更高。此外,纵向研究追踪了自闭症症状随时间的发展,揭示了可以为早期干预和诊断提供信息的模式和轨迹。总体而言,这些研究有助于我们了解自闭症的复杂性和多面性,强调需要进一步研究其病因、病程和治疗。研究调查了有自闭症谱系障碍 (ASD) 风险的婴儿的早期运动能力和行为。这些研究旨在确定婴儿时期自闭症的潜在标志或指标,希望它们可以用作预测指标或后期诊断的预测指标。2019 年发表的一项研究发现,患自闭症风险较高的婴儿与风险较低的婴儿相比,表现出不同的运动能力。2012 年发表的另一项研究发现,婴儿的头部滞后与患自闭症的风险增加之间存在相关性。研究人员还探讨了注意力、社交参与和视觉处理在有自闭症风险的婴儿中的作用。例如,一项研究发现,后来被诊断为自闭症的婴儿早在 6 个月大时就表现出对社交场景的自发注意力下降。另一项研究发现,这些婴儿在受到干扰时不太可能与自己的面部互动。此外,研究还检查了有自闭症风险的婴儿对言语提示和听觉刺激的反应。2017 年发表的一项研究发现,这些婴儿对名字识别的反应与没有自闭症的婴儿不同。最近的研究集中于婴儿期的大脑发育,包括白质微结构、皮层下脑功能和皮层厚度。这些研究旨在确定 ASD 的潜在生物标记或了解潜在的神经机制。总体而言,这些研究表明,早期运动能力、注意力、社交参与和视觉处理可能是婴儿期 ASD 风险的重要指标。需要进一步研究才能充分了解这些因素与 ASD 发展之间的关系。研究表明,婴儿的白质微结构发育与认知能力密切相关。研究使用基于束的分析和功能连接映射等技术,研究了从出生到 2 岁期间大脑结构和功能的变化。一项研究发现,0-24 个月大婴儿的白质结构变化与 24 个月大婴儿的认知能力提高有关(Gao 等人,2015 年)。另一项研究发现,在 6 至 18 个月大婴儿出现自闭症样症状时,婴儿在 12 个月大时某些大脑区域的白质完整性会降低,这预示着 24 个月大婴儿的诊断结果会更准确(Emerson 等人,2017 年)。其他研究人员使用 MRI 扫描检查了患有自闭症谱系障碍 (ASD) 的婴儿的大脑,发现婴儿的大脑结构存在显著差异,包括某些区域的大脑尺寸增大(Piven 等人,1992 年;Courchesne 等人,2001 年)。一项较新的研究使用大量高风险婴儿,确定了自闭症的早期生物标志物,例如大脑区域间功能连接减少(Hazlett 等人,2017 年)。这些发现表明,早期生活经历和遗传倾向可以影响自闭症儿童的大脑发育。需要进一步研究以了解推动这些变化的潜在机制并制定有效的干预措施。注意:我将参考文献压缩为较短的格式,同时保留基本信息。如果您希望我扩展任何特定参考文献或提供更多详细信息,请告诉我!一系列研究调查了自闭症谱系障碍 (ASD) 患者从出生到 2 岁及以后的大脑发育和结构。该研究使用磁共振成像 (MRI) 和头围测量来检查自闭症儿童的大脑大小和形状。研究发现,在幼儿时期,大脑增大与自闭症男孩的退化有关。此外,后来患上自闭症的个体的皮质表面积在 2 岁之前增加。纵向 MRI 研究表明,自闭症患者的皮质发育持续到儿童早期。其他研究表明,2-3 岁的幼儿就存在脑成像异常,这表明自闭症可能是一种早期神经发育障碍。一些研究发现,一部分患有自闭症的男孩的表面积增加,但皮质厚度没有增加,而其他研究则使用基于表面的形态测量法来绘制患有自闭症的学龄前儿童的皮质解剖图。总体而言,研究表明,自闭症患者的大脑结构和发育从儿童早期开始就会发生改变。自闭症谱系障碍 (ASD) 的研究表明,大脑结构的变化,特别是皮质厚度的变化,可能与自闭症有关。研究发现,与没有自闭症的人相比,自闭症患者的皮质厚度往往会发生变化。然而,这些变化的程度和性质在不同的发育阶段会有所不同。一些研究表明,患有自闭症的儿童表现出额叶皮质褶皱增加,而年龄较大的青少年和成年人则表现出额叶皮质褶皱减少。此外,研究发现,患有自闭症的个体经常表现出脑沟大小和形状异常,这可能与社交沟通障碍有关。杏仁核是参与情绪处理的区域,也与自闭症有关。研究表明,患有自闭症的个体往往比没有自闭症的个体拥有更大的杏仁核,尤其是在幼儿和幼儿中。然而,杏仁核大小和自闭症行为症状之间的关系很复杂,受各种因素的影响。纵向研究为自闭症大脑变化的发展和进展提供了宝贵的见解。例如,一项研究发现,自闭症儿童的杏仁核体积随着年龄的增长而增加,而另一项研究发现,联合注意力技能与杏仁核体积的变化有关。脆性 X 综合征的研究强调了自闭症的异质性,脆性 X 综合征与自闭症有一些相似之处,但也表现出明显的大脑差异。总体而言,研究结果表明,大脑结构和功能在理解自闭症方面发挥着重要作用,需要进一步研究来阐明大脑变化与自闭症行为症状之间的复杂关系。一些发表在知名期刊上的重要研究包括:* Wolff 等人(2014 年)- 神经发育障碍:通过发展研究加速自闭症的进展。* Libero 等人(2018 年)- 自闭症谱系障碍年轻男孩局部脑回指数的纵向研究。* Williams 等人(2012 年)- 自闭症和阅读障碍皮质复杂性的球谐分析。* Kohli 等人(2019 年)- 自闭症谱系障碍儿童的局部皮质脑回增加,但青少年的局部皮质脑回迅速减少。这些研究表明,人们正在努力了解自闭症的神经基础,并开发有效的干预措施来支持患有这种疾病的人。研究调查了与自闭症谱系障碍 (ASD) 相关的大脑结构和发育变化。研究发现,患有自闭症的儿童,尤其是 2-5 岁的儿童,尾状核发育异常,而尾状核与重复行为有关。此外,由于家庭因素而患自闭症风险较高的婴儿被发现存在大脑解剖结构差异,包括皮层下和小脑区域,这预示着以后重复行为的出现。纵向研究还表明,患有自闭症的幼儿随着时间的推移,胼胝体形态会发生变化。这些变化可能与自闭症相关症状的发展有关,例如社交互动受损和沟通困难。此外,研究强调了小脑在自闭症中的潜在作用,几项研究表明自闭症患者的小脑体积和结构发生了改变。小脑参与运动控制、学习和情绪调节,可能导致自闭症中观察到的认知和行为症状。总体而言,这些发现表明,大脑发育和解剖结构的早期变化可能与自闭症症状的出现有关,特别是那些与重复行为和社交沟通困难相关的症状。本文讨论了一系列关于自闭症谱系障碍 (ASD) 儿童大脑发育的研究。该研究重点关注了自闭症儿童与非自闭症儿童相比,大脑中白质纤维和连接的发育情况。一项研究发现,自闭症儿童在幼儿期白质成熟速度加快。另一项研究发现自闭症儿童的白质完整性存在异常。第三项研究表明,后来被诊断患有自闭症的幼儿颞胼胝体纤维表现出多种结构异常。其他研究使用弥散张量成像 (DTI) 来研究自闭症儿童白质纤维和连接的发育情况。一项研究发现,6-24 个月之间,自闭症婴儿与非自闭症婴儿的白质纤维束发育存在差异。另一项研究发现,自闭症幼儿的白质连接异常,包括额叶可能存在轴突过度连接。总体而言,这些研究表明,自闭症儿童的白质纤维和连接发育可能存在异常,这可能与该疾病特有的社交和认知缺陷有关。研究发现,自闭症谱系障碍 (ASD) 患者的网络效率低下早在 24 个月大时就存在,这种现象可能会持续存在并随着时间的推移发展成更严重的症状。研究表明,有患自闭症风险的婴儿在 6-7 个月大时就会表现出异常的神经回路和白质微结构,尤其是在负责语言处理和社交互动的区域。此外,研究还发现,自闭症患者的大脑中与语言处理相关的脑区侧化往往会发生改变,这会影响他们处理和理解语言的能力。这种语言障碍被认为是自闭症早期出现的根本特征。此外,研究表明,自闭症幼儿的神经同步模式被破坏,这可能导致自闭症特有的社交和沟通障碍的发展。研究还探讨了自闭症幼儿的大脑功能与社交行为之间的关系。研究发现,与社交和感觉运动缺陷相关的神经回路功能连接异常可以预测自闭症的后期症状。最后,研究检查了患有自闭症的学龄前男孩语言变异的神经相关性,发现非典型语言处理模式与患自闭症的风险更高有关。总体而言,这些研究表明,大脑结构和功能的早期异常可能导致自闭症症状的发展,并强调早期诊断和干预的必要性。研究表明,患有自闭症谱系障碍 (ASD) 的儿童表现出大脑连接和发育中断,特别是在杏仁核和胼胝体等区域。研究还发现白质纤维束发育存在差异,这可能导致自闭症的发展。有患自闭症风险的婴儿的大脑功能和结构发生了改变,包括白质和胼胝体的变化。这些发现表明,早期干预可能有助于预防或减轻自闭症的影响。此外,研究表明,更广泛的自闭症表型早在婴儿期就可能出现,这表明自闭症是一种复杂的特征,不能仅仅归因于遗传因素。此外,对人类大脑发育的研究揭示了神经干细胞和祖细胞在大脑皮层形成中的作用。研究还表明,进化过程中的大脑皮层扩张可能与自闭症的病因有关。总的来说,这些发现强调了早期发现和干预自闭症风险患者的重要性,以及进一步研究这种复杂疾病的潜在机制和原因的必要性。注意:我在改写过程中进行了一些创造性的改动,使其更易读、更简洁,同时保持了原文的整体含义和本质。对特发性自闭症患者神经细胞的研究表明,其增殖和网络发生了改变(Marchetto 等人,2017 年)。此外,研究发现,自闭症谱系障碍中存在 FOXG1 依赖的 GABA/谷氨酸神经元分化失调(Mariani 等人,2015 年)。此外,病理性启动与自闭症受试者衍生神经元的发育基因网络异时性有关(Schafer 等人,2019 年)。遗传学研究已确定了与自闭症有关的几个关键基因,包括 PTEN,它调节小鼠的神经元树突和社交互动(Kwon 等人,2006 年)。破坏性 CHD8 突变也已被证明可在发育早期定义自闭症亚型(Bernier 等人,2014 年)。已经研究了来自自闭症谱系障碍遗传模型的人类诱导多能干细胞衍生神经元的细胞表型,揭示了与典型对照细胞相比的细胞差异(Deshpande 等人,2017 年)。在 16p11.2 缺失和重复变异的携带者中也发现了相反的大脑差异(Qureshi 等人,2014 年)。研究探索了小鼠大脑皮层上层神经元与自闭症样特征之间的关系,揭示了这些神经元的过量生产导致自闭症行为(Fang 等人,2014 年)。改变的大脑皮层基因表达、大脑过度生长和功能过度连接也与小鼠的 Chd8 单倍体不足有关(Suetterlin 等人,2018 年)。对发育突触修剪的研究揭示了 LTD 样分子通路在此过程中的作用,对自闭症研究具有潜在意义(Piochon 等人,2016 年)。局部皮质回路的关键期可塑性也得到了探索,强调了这一时间窗口对大脑正常发育和功能的重要性(Hensch,2005)。已发现导致综合征性自闭症的突变定义了突触病理生理学轴,这对我们理解自闭症的潜在机制具有重要意义(Auerbach 等人,2011)。研究人员发现,综合征性和非综合征性自闭症啮齿动物模型中存在共同的突触病理生理学。研究还表明,在携带与自闭症相关的拷贝数变异的小鼠中,小脑可塑性和运动学习能力受损。此外,有自闭症风险的婴儿的白质微结构发生了改变,表明早期大脑发育发生了变化。已发现脑脊液 (CSF) 在神经祖细胞增殖中起着至关重要的作用,可能参与自闭症的早期大脑发育。 CSF 还提供了清除间质溶质(包括淀粉样蛋白 β)的途径。髓鞘形成缺陷在综合征型和特发性自闭症谱系障碍 (ASD) 中很常见。Pten 的体质性错误定位与少突胶质细胞的早熟和 ASD 模型中的异常髓鞘形成有关。前额叶轴突的变化可能会破坏自闭症中的网络,表明连接性发生了改变。活动依赖性髓鞘形成和髓鞘形成神经胶质细胞上的非突触连接促进电活性轴突的优先髓鞘形成。最后,几项研究已经确定了自闭症谱系障碍 (ASD) 的常见遗传风险变异,强调了了解这种疾病背后的复杂遗传学的重要性。这一系列参考文献涉及自闭症谱系障碍 (ASD) 及其早期检测和预测。这些论文探讨了各个方面,包括:* ASD 的生物学,从细胞增殖到临床表型 * 父母的担忧可以预测以后的自闭症诊断 * 18 个月的标记可以预测自闭症儿童的弟弟妹妹以后的结果 * 幼儿自闭症的筛查工具 * 对疑似患有自闭症的幼儿的临床评估和管理 * 使用深度学习和机器学习算法研究精神和神经疾病(包括自闭症)的神经影像相关性 此外,参考文献还涉及: * 深度学习在婴儿脑部 MRI 分析中的作用 * 解决不平衡数据集和改进预测模型的技术 * 从出生到婴儿期自闭症患者的大脑和行为发展 * 预测是人类认知神经科学对人道主义和务实应用的贡献 * 跟踪精神病的维度和分类特征的个体特定功能连接标记 总体而言,这些参考文献为自闭症的早期发现和预测以及机器学习和深度学习算法在该领域的应用提供了见解。最近的研究探索了精神神经影像学在临床环境中的应用,即心理放射学。该领域已显示出利用磁共振成像 (MRI) 和放射组学分析检测精神分裂症和注意力缺陷多动障碍 (ADHD) 的前景。具体而言,研究重点是通过分析大脑图像和基于图形的指标来提高 ADHD 的诊断准确性。研究还检验了对患有自闭症谱系障碍的幼儿进行早期干预的有效性,包括父母介导的疗法和行为干预。此外,人们对精准医疗的兴趣日益浓厚,精准医疗旨在根据患者独特的基因特征为其量身定制治疗方案。另一个研究领域涉及了解自闭症的神经相关性,一些研究表明,出生时的白质连接组可以预测成年后的认知能力。此外,脑成像和机器学习的进步使研究人员能够开发出分析大脑网络和预测神经发育结果的新工具。这些发现对一系列神经和精神疾病的早期诊断、治疗和干预策略具有潜在意义。提到的一些关键研究包括:* Lei 等人。 (2019):研究了全脑图像、全连接组功能连接和基于图形的指标在检测精神分裂症方面的相对诊断价值。 * Port JD (2018):提出使用 MRI 成像和放射组学分析来诊断 ADHD。 * Collins & Varmus (2015):提出了一项关于精准医疗的新举措。 * Dawson 等人 (2010):对患有自闭症的幼儿进行了一项早期干预的随机对照试验,称为早期丹佛模式。 * Kasari 等人 (2015):评估了家长干预对自闭症幼儿的比较效果。这些研究表明,我们正在不断努力提高对精神神经影像学及其在临床环境中的应用的理解。以下文章讨论了自闭症谱系障碍 (ASD) 研究的各个方面,包括诊断、认知特征、大脑功能和遗传因素。这些研究探讨了理解 ASD 的不同方法,例如使用机器学习算法来识别认知特征的亚型,分析静息状态功能网络来识别大脑系统的个体特定特征。文章还讨论了考虑 ASD 异质性的重要性,一些研究侧重于父母认知和行为特征在塑造临床变异性方面的作用。其他研究则探讨了早期运动迟缓与后来诊断 ASD 或表达性语言困难之间的关系。这项研究强调了精准医疗方法的必要性,以了解 ASD 的复杂原因,重点是开发更有效的诊断工具和治疗方法。研究表明,常见的遗传变异、多基因风险和罕见基因突变的附加效应都会增加患 ASD 的风险。 2012 年和 2016 年的两篇研究论文研究了有自闭症风险的婴儿的沟通延迟。这些研究发现,早期运动技能可以预测自闭症谱系障碍儿童的语言发育。2019 年发表的另一项研究讨论了基于干细胞的脑器官的出现,这可能有助于理解发育障碍。日本的一系列病例使用视频脑电图监测研究了四名 8 至 14 个月大婴儿的颤抖发作 (SA)。发作的特征是凝视、紧张和颤抖,通常在清醒时成群发生,没有意识障碍。不寻常的动作包括罕见的眼球反张姿势和眼球偏斜。这些发作通常由换尿布或喂食时间引发,在 2-3 岁时会消退或显着改善。颤抖发作可能会被误诊为癫痫;然而,在大多数情况下,它们被认为是一种可以消退的良性疾病。另一种需要考虑的情况是由于镁营养缺乏导致的婴儿震颤综合征,这种综合征会导致快速震颤,在睡眠期间会消失,需要镁治疗,但可能无法纠正精神运动发育迟缓。这些研究探讨了理解自闭症的不同方法,例如使用机器学习算法来识别认知特征的亚型,分析静息状态功能网络来识别大脑系统的个体特定特征。文章还讨论了考虑自闭症异质性的重要性,一些研究侧重于父母的认知和行为特征在塑造临床变异性方面的作用。另一些研究则探讨了早期运动迟缓与后来诊断自闭症或表达性语言困难之间的关系。这项研究强调了精准医疗方法的必要性,以了解自闭症的复杂病因,重点是开发更有效的诊断工具和治疗方法。研究表明,常见的遗传变异、多基因风险和罕见基因突变的附加效应都会增加患自闭症的风险。2012 年和 2016 年的两篇研究论文研究了有自闭症风险的婴儿的沟通延迟。这些研究发现,早期运动技能可以预测自闭症谱系障碍儿童的语言发展。2019 年发表的另一项研究讨论了基于干细胞的脑类器官的出现,这可能有助于了解发育障碍。日本的一系列病例使用视频脑电图监测研究了 4 名 8 至 14 个月大婴儿的颤抖发作 (SA)。发作的特征是凝视、紧张和颤抖,通常在清醒时成群发生,没有意识障碍。异常动作包括罕见的反张姿势和眼球偏斜。发作通常由换尿布或喂奶时间引发,在 2-3 岁时会消退或显着改善。颤抖发作可能被误诊为癫痫;然而,在大多数情况下,它们被认为是一种良性疾病,可以得到解决。另一种需要考虑的疾病是由于镁营养缺乏引起的婴儿震颤综合征,它会导致快速震颤,在睡眠中消失,需要镁治疗,但可能无法纠正精神运动发育迟缓。这些研究探讨了理解自闭症的不同方法,例如使用机器学习算法来识别认知特征的亚型,分析静息状态功能网络来识别大脑系统的个体特定特征。文章还讨论了考虑自闭症异质性的重要性,一些研究侧重于父母的认知和行为特征在塑造临床变异性方面的作用。另一些研究则探讨了早期运动迟缓与后来诊断自闭症或表达性语言困难之间的关系。这项研究强调了精准医疗方法的必要性,以了解自闭症的复杂病因,重点是开发更有效的诊断工具和治疗方法。研究表明,常见的遗传变异、多基因风险和罕见基因突变的附加效应都会增加患自闭症的风险。2012 年和 2016 年的两篇研究论文研究了有自闭症风险的婴儿的沟通延迟。这些研究发现,早期运动技能可以预测自闭症谱系障碍儿童的语言发展。2019 年发表的另一项研究讨论了基于干细胞的脑类器官的出现,这可能有助于了解发育障碍。日本的一系列病例使用视频脑电图监测研究了 4 名 8 至 14 个月大婴儿的颤抖发作 (SA)。发作的特征是凝视、紧张和颤抖,通常在清醒时成群发生,没有意识障碍。异常动作包括罕见的反张姿势和眼球偏斜。发作通常由换尿布或喂奶时间引发,在 2-3 岁时会消退或显着改善。颤抖发作可能被误诊为癫痫;然而,在大多数情况下,它们被认为是一种良性疾病,可以得到解决。另一种需要考虑的疾病是由于镁营养缺乏引起的婴儿震颤综合征,它会导致快速震颤,在睡眠中消失,需要镁治疗,但可能无法纠正精神运动发育迟缓。以及罕见基因突变的附加效应都会增加患自闭症的风险。2012 年和 2016 年的两篇研究论文研究了有自闭症风险的婴儿的沟通延迟。这些研究发现,早期运动技能可以预测自闭症谱系障碍儿童的语言发展。2019 年发表的另一项研究讨论了基于干细胞的脑器官的出现,这可能有助于理解发育障碍。日本的一系列病例使用视频脑电图监测研究了四名 8 至 14 个月大婴儿的颤抖发作 (SA)。发作的特征是凝视、紧张和颤抖,通常在清醒时成群发生,没有意识障碍。不寻常的动作包括罕见的反张姿势和眼球偏斜。这些发作通常由换尿布或喂食时间引发,在 2-3 岁时会消退或显着改善。颤抖发作可能会被误诊为癫痫;然而,在大多数情况下,它们被认为是一种良性疾病,可以得到解决。另一种需要考虑的情况是由于镁营养缺乏导致的婴儿震颤综合征,这种综合征会导致快速震颤,在睡眠期间会消失,需要镁治疗,但可能无法纠正精神运动发育迟缓。以及罕见基因突变的附加效应都会增加患自闭症的风险。2012 年和 2016 年的两篇研究论文研究了有自闭症风险的婴儿的沟通延迟。这些研究发现,早期运动技能可以预测自闭症谱系障碍儿童的语言发展。2019 年发表的另一项研究讨论了基于干细胞的脑器官的出现,这可能有助于理解发育障碍。日本的一系列病例使用视频脑电图监测研究了四名 8 至 14 个月大婴儿的颤抖发作 (SA)。发作的特征是凝视、紧张和颤抖,通常在清醒时成群发生,没有意识障碍。不寻常的动作包括罕见的反张姿势和眼球偏斜。这些发作通常由换尿布或喂食时间引发,在 2-3 岁时会消退或显着改善。颤抖发作可能会被误诊为癫痫;然而,在大多数情况下,它们被认为是一种良性疾病,可以得到解决。另一种需要考虑的情况是由于镁营养缺乏导致的婴儿震颤综合征,这种综合征会导致快速震颤,在睡眠期间会消失,需要镁治疗,但可能无法纠正精神运动发育迟缓。