XiaoMi-AI文件搜索系统
World File Search System文章
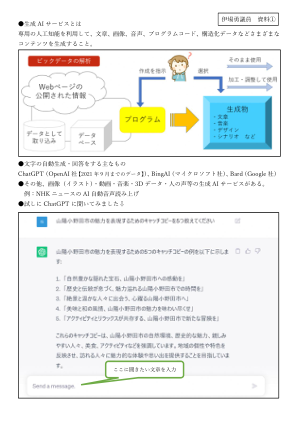

引入利用AI(人工智能)的文本分析技术
从客户保护角度,本行分支机构销售人员在分支机构推介、销售金融产品或拜访客户时,会记录与客户的互动情况,本行总行合规部门则会监控客户在介绍金融产品时所作说明的恰当性,以及对产品和投资风险的了解程度。然而,目视检查存在判断标准不一致、数据量过大导致问题发现延迟等问题,因此只能进行局部检查。

文章
葡萄是全球公认的具有重要经济价值的果树。在葡萄品种中,汤普森无核葡萄对鲜食和酿酒、干燥和榨汁的广泛应用具有至关重要的影响。该品种是葡萄基因改造最有效的基因型之一。然而,缺乏高质量的基因组阻碍了有效的育种工作。在这里,我们展示了汤普森无核葡萄的高质量参考基因组,其中所有 19 条染色体都表示为 19 个连续序列(N50 = 27.1 Mb),没有间隙,并且预测了所有端粒和着丝粒。与之前的组装(TSv1 版本)相比,新组装包含额外的 31.5 Mb 高质量测序数据,注释了总共 30 397 个蛋白质编码基因。我们还进行了细致的分析,以确定汤普森无核葡萄和两种以抗病性而闻名的野生葡萄品种中的核苷酸结合亮氨酸富集重复基因 (NLR)。我们的分析表明,汤普森无核葡萄中两种类型的 NLR(TIR-NB-LRR (TNL) 和 CC-NB-LRR (CNL))的数量显著减少,这可能导致其对许多真菌疾病(如白粉病)敏感,而第三种类型的 NLR(RPW8(抗白粉病 8)-NB-LRR (RNL))的数量增加。随后,转录组分析表明,在白粉病感染期间 NLR 显著富集,强调了这些元素在葡萄树防御白粉病中的关键作用。高质量汤普森无籽参考基因组的成功组装对葡萄基因组学研究做出了重要贡献,深入了解了无籽、抗病性和颜色性状的重要性,这些数据可用于促进葡萄分子育种工作。

文章
质子陶瓷电化学电池(PCEC)是一种很有前途的固态能量转换装置,它能够在中温度下实现电能和氢能之间的能量转换。通过PCEC技术实现化学能和电能之间的快速转换将有助于应对能源存储的巨大挑战。为了实现制氢和发电之间的高效可逆操作,在保持持久运行的同时提高氧电极的水氧化和氧还原活性是早期的技术机会之一。在本研究中,A位缺陷层状钙钛矿(PrBa 0.8 Ca 0.2 ) 0.95 Co 2 O 6-δ已被开发为PCEC中的氧电极,其具有优异的电化学性能。电解电流密度在 1.3 V 时高达 -0.72 A cm -2,在电解和燃料电池模式下分别在 600 ○ C 下获得 0.540 W cm -2 的峰值功率密度。采用新电极的 PCEC 在两种操作模式下均表现出良好的耐久性,在 160 小时内没有明显的性能下降。电解和燃料电池模式之间的可逆性也得到成功展示。

文章
直接生长技术用于合成几个宏观工程师(MMS),该技术采用可逆的添加 - 碎片链转移(RAFT)聚合通过直接从诺尔伯伦官能官能化的链转移剂(CTA)生长。我们的目的是研究由四种单体中不同单体转化值在不同的单体转化值下通过组合(即耦合)终止的双生烯基物质的形成:苯乙烯,丁烷,丁酸 - 丁基 - 丙烯酸丙烯酸酯,甲基丙烯酸甲酯和N-丙烯酸甲基丙烯酸甲酯和甲基丙烯酰胺。使用Grubbs 3 Rd Generation催化剂(G3)以mm:G3比为100:1的Grubbs对这些MM的开环式聚合(ROMP),导致瓶颈聚合物的形成。通过尺寸排斥色谱分析(SEC)表明,高摩尔质量肩部的高巨大强度归因于掺入这些双苯二烯基物质以产生二聚体或高阶瓶洗聚合物寡聚物。The monomer type in the RAFT step heavily influenced the amount of these bottlebrush polymer dimers and oligomers, as did the monomer conversion value in the RAFT step: We found that the ROMP of polystyrene MMs with a target backbone degree of polymerization of 100 produced detectable coupling at 20% monomer conversion in the RAFT step, while it took 80% monomer conversion to observe在聚(Tert-丁基丙烯酸酯)MMS中耦合。我们没有检测到聚(甲基丙烯酸甲酯)MMS中的耦合,但是SEC峰扩大并增加了分散性的增加,这表明存在隔离活性烷烃链链,这是由抗倍率所产生的。最后,即使在木筏步骤中达到90%的单体转换时,poly(n-丙烯酰基)MMS也没有显示出瓶洗瓶装聚合物的可检测耦合。这些结果突出了单体选择和筏聚合条件在制作摩擦术的MMS中的重要性,以制造定义明确的瓶洗聚合物。

文章
共晶工程作为一种生产具有有趣特性的新材料的有前途的方法正受到越来越多的关注,正在进行的研究正在开发可靠的共结晶设计规则。1 2 3 4 组成分子(本文称为构造子)5 的大小和形状是控制晶格排列的重要因素,同时还影响固态填充产生的紧密分子间相互作用的强度和方向性。6 7 原则上,当所有构造子都是具有明确形状的刚性分子时,预测可能的共晶格填充相对容易。如果部分或全部构造子都是柔性分子,则共晶格预测变得更具挑战性。6 7 8 在这种情况下,最简单的概念方法是假设所有柔性构造子都采用其最低能量构象。然而,这种范式忽略了高能构象中的柔性构造子可能由于功能组定位改善而允许更有利的固态填充的可能性。换句话说,增加有利的分子间相互作用的数量可以抵消构造子采用高能分子构象时产生的能量损失。6 9

文章
本研究研究了脉冲CF 3 I/C 4 F 8 /Ar/O 2 电感耦合等离子体用于低k刻蚀,研究了C 4 F 8 /Ar/O 2 中添加CF 3 I对等离子体特性和低k材料刻蚀特性的影响。随着混合气体中CF 3 I/(CF 3 I + C 4 F 8 )比例的增加,等离子体中CF 3 自由基增多,CF 2 自由基减少,其中CF 3 自由基和CF 2 自由基分别与刻蚀和聚合有关。因此,SiCOH的刻蚀速率随CF 3 I比例的增加而增大。然而,当CF 3 I比例为0.5时,等离子体中的CF 2 /F通量比和聚合物层上的C/F比最高,因此对非晶碳层和光刻胶的刻蚀选择性在比例为0.5时最高。 SiCOH 损伤随 CF 3 I 比率的增加而减小,并且 SiCOH 损伤似乎非常低,特别是当 CF 3 I 比率≥0.5 时,Si–CH 3 键损失低、F 渗透低、表面粗糙度低。因此,与仅使用 C 4 F 8 /Ar/O 2 气体混合物相比,将 50% CF 3 I 混入 C 4 F 8 /Ar/O 2 气体混合物中不仅可以产生相对于掩模材料的高蚀刻选择性,而且还可能减少蚀刻损伤。

文章
了解在复杂环境(例如,人体,土壤和海洋环境)中基于金属的纳米颗粒与生物系统之间的相互作用仍然具有挑战性,尤其是在单细胞和纳米级水平上。捕获这些相互作用的动力学,例如金属分布,纳米颗粒的生长和降解,在其本地状态(体内)特别困难。在这里,我们证明了使用基于同步加速器的X射线纳米仪扫描荧光显微镜与液体细胞环境相结合的水合磁铁矿磁性磁性细菌中铁含量的直接测量。除了X射线荧光成像外,我们还使用X射线吸收光谱从液体中的单个细菌中收集了铁化学物种形成信息。对于原位生物矿化研究,我们开发了一种微流体装置,以在X射线束下的几个小时内跟踪磁铁矿纳米颗粒形成。这种方法突出了液体细胞设置中X射线荧光显微镜的潜力,以在单细胞水平上为生物过程提供元素和化学见解。将X射线纳米膜技术与液体细胞设备相结合,可以在同步加速器上进行生物学环境中的金属实验。

文章
大学里尔,inria,CNRS,中央里尔,UMR 9189 - 法国克里斯塔尔(P Saux MSC,J Teigny Beng,T Soumphonphakdy MSC); University,CNRS,Inria,Central Lille,UMR 9189 - Cristal,Lille,France(P教授P Preus博士);大学里尔大学,Inserm,Chu Lille,Pastere Lille,U1190-EGID,Lille,法国(P Bavin Phd,V Caiazo MD,R Caiazo MD教授,MD教授,F Pattou Pattou MD,F Pattou f Pattou Pattou Md Phd教授);荷兰语Obesia,House Ter Heide,荷兰(雅各布斯MSC,D Jacobs BSC,V Monelle PhD);新加坡新加坡综合医院的部门或内部科学部(P C C Lee MBCHB);新加坡新加坡综合医院的部门或胃中睾丸和减肥手术,分区或手术(C H Lircs);系或分子和临床医学,大学或医学,大学或哥德堡的Shlgrenska学院,哥德堡,瑞典博士,L Carlsson MD教授,P-A Svensson Pr.
