XiaoMi-AI文件搜索系统
World File Search System旁系

四个BNITPK基因在四倍体油籽中的基因编辑导致种子中的植酸显着降低
甘蓝纳普斯的摘要商业化。l(油籽)餐正在越来越关注。植酸(PA)是植物中磷的主要来源,但由于人类对基本矿物质吸收的不利影响,对包括人类在内的单胃动物被认为是抗营养。未消化的PA会导致富营养化,这可能威胁着水生生命。pa在油料强奸的成熟种子中占2-5%,并通过涉及多种酶的复杂途径合成。隐性性状的多倍体繁殖多倍体具有挑战性,因为基因功能由几个旁系同源物编码。基因冗余通常需要淘汰几个基因副本以研究其潜在效果。因此,我们采用了CRISPR-Cas9诱变来淘汰BNITPK的三个功能旁系同源物。我们获得了低pa突变体,而在低芥酸菜籽级春季品种海丁中,游离磷的增加。这些突变体可以标志着菜籽繁殖的重要里程碑,蛋白质价值增加,对油含量没有不利影响。

文章 以 CRISPR/Cas9 为基础的靶向敲除水稻直系同源物 TILLER ANGLE CONTROL 1 (TAC1) 可诱导杨树直立叶片习性和枝条生长
摘要:金字塔形、直立或直立生长的植物形态的特点是枝条和叶子的分枝角度较窄。直立叶子和枝条习性的优势可能是光线更有效地穿透较低的冠层。已经报道了包括桃树在内的各种树种的金字塔基因型。旁系同源水稻直系同源物 TILLER ANGLE CONTROL 1 (TAC1) 被认为是负责直立生长的基因。然而,对于任何金字塔树种基因型,尚未真正证明 TAC1 基因的敲除突变会导致植物金字塔形生长。通过计算机分析,我们在 P. trichocarpa 基因组中发现了一个假定的水稻 TAC1 直系同源物(Potri.014G102600,“TAC-14”)及其旁系同源物(Potri.002G175300,“TAC-2”)。通过应用转基因 CRISPR/Cas9 方法成功敲除 P. × canescens 克隆 INRA 717-1B4 中的两个假定的 PcTAC1 直系同源物。在温室中对突变体进行了为期三年的分子分析和表型分析。我们的结果表明,“TAC-14”的纯合敲除足以诱导 P. × canescens 中的金字塔形植物生长。如果在短轮伐期林(SRC)上种植多达两倍的金字塔树种,那么可以提高木材产量,无需任何育种,只需增加默认田地面积上的树木数量即可。

数字化如何改变铁路基础设施
ETCS 上的 ATO:工作原理 ATO 轨旁系统收集线路上的静态和动态数据以及交通管理系统提供的时间表,并将其传输到车辆。根据基础设施、轨道和时间表的可用数据,车载系统可随时计算最佳行程曲线,并在列车自动运行时控制车辆的牵引和制动系统。机车司机通过显示屏持续了解所有程序。因此,能耗降低,交通流量和容量显著提高。由于自动制动,ETCS 上的 ATO 缩短了制动距离,从而大大改善了间隔时间。

用crispr/cas9
摘要:用CRISPR/CAS9靶向基因组是引入突变和产生敲除效应的流行方法。但是,目前可以提供有关基本基因诱变的有限信息。这项研究研究了CRISPR/CAS9在靶向水稻必不可少的基因中的效率:雷帕霉素的单胎靶标(ostor)的单胎靶标和可可糖非发酵-1(SNF1)相关的激酶1的三个旁系同源物(OSSNRK1α),OSSNRK1α,OSSNRK和OSSNRK1 C.组成型表达的CRISPR/CAS9可有效地在Ostor和OssnRK1α基因中产生突变,但是诱导的CRISPR/CAS9未能产生可检测的突变。术的诱变速率相对较低,只能靶向骨的激酶结构域,而热区域的突变则无法恢复。ossnrk1α旁系同源物可以以较高的速率作为目标;但是,在> 50%的主要突变体中观察到不育或早期衰老。此外,具有高序列同源性的OSSNRK1αB和OSSNRK1αC可以同时靶向以产生双突变剂。此外,尽管在幸存的突变体中发现了有限类型的突变,但恢复的线表现出功能丧失或敲低的TOR或SNRK1表型。总的来说,我们的数据表明,CRISPR/CAS9可以创建这些基本基因中的突变,以促进对其在大米中植物发育和环境反应中的作用的研究。

一个共享的古老增强子元件差异地调节果蝇腿部发育过程中的 bric-a-brac 串联基因重复
基因复制和转录增强子的出现/修饰被认为对动物进化过程中表型创新做出了巨大贡献。尽管如此,人们对基因复制后增强子如何进化以及调控信息如何在复制基因之间重新连接知之甚少。果蝇 bric-a-brac (bab) 复合体由串联旁系同源基因 bab1 和 bab2 组成,为解决这些问题提供了范例。我们之前描述了一种调节发育足中 bab2 表达的基因间增强子 (名为 LAE)。我们在此显示直接与 LAE 结合的 bab2 调节子也控制跗骨细胞中的 bab1 表达。通过 CRISPR/Cas9 介导的基因组编辑切除 LAE 表明,这种增强子似乎参与了 bab1 和 bab2 在腿部组织中共表达,但并不是严格必需的。相反,LAE 增强子对于沿近端-远端足轴的旁系同源物特异性 bab2 表达至关重要。染色质特征和表型挽救实验表明,LAE 功能部分冗余,腿特异性调控信息与 bab1 转录单元重叠。系统基因组学分析表明 (i) bab 复合体起源于 Cyclorrhapha dipteran 亚系早期祖先单基因的复制,以及 (ii) LAE 序列在 Brachycera 亚目中很早就已进化固定,因此早于基因复制事件。这项工作为增强子提供了新的见解,特别是关于它们的出现、维持和进化过程中的功能多样化。
小鼠结核病感染过程中对IRGM蛋白的差异需求
摘要 结核分枝杆菌 ( Mtb ) 是一种仅寄居在人类宿主中的细菌,并且是全球传染病发病率和死亡率的主要原因。宿主对 Mtb 感染的保护依赖于免疫相关 GTPase 进化枝 M (IRGM) 蛋白的功能。人类 IRGM 的多态性与对分枝杆菌病的易感性改变有关,并且人类 IRGM 促进 Mtb 进入降解的自噬溶酶体。在三种鼠 IRGM 直系同源物中,Irgm1 被认为是在培养的巨噬细胞和体内 Mtb 感染期间宿主保护所必需的。然而,旁系同源鼠 Irgm 基因 Irgm2 和 Irgm3 是否在宿主防御 Mtb 中发挥作用或在 Mtb 感染期间与 Irgm1 表现出功能关系仍未确定。这里我们报告 Irgm1 2 / 2 小鼠确实对结核分枝杆菌气溶胶感染极为敏感,而额外删除旁系同源物 Irgm3 基因可恢复 Irgm1 缺陷动物对结核分枝杆菌感染的保护性免疫力。缺乏所有三个 Irgm 基因(泛 Irgm 2 / 2 )的小鼠的特点是感染后 5 和 24 周肺细胞因子谱发生变化,但可控制病情直至感染的晚期阶段,此时泛 Irgm 2 / 2 小鼠与野生型小鼠相比死亡率增加。总之,我们的数据表明 Irgm 亚型之间平衡的破坏对结核分枝杆菌感染宿主的危害比 Irgm 介导的宿主防御完全丧失更大,这一概念也需要在与 IRGM 多态性相关的人类结核分枝杆菌易感性的背景下考虑。

CRISPR-Cas9 靶向 eIF4E1 基因可扩大 Solanum tuberosum L. cv. Desirée 的马铃薯 Y 病毒抗性谱
翻译起始因子,特别是 eIF4E 家族,是许多植物物种对马铃薯 Y 病毒组隐性抗性的主要来源。然而,在马铃薯 (Solanum tuberosum L.) 种质中尚未鉴定出 eIF4E 介导的对该病毒属的抗性。与番茄一样,马铃薯 eIF4E 基因家族由 eIF4E1、其旁系同源物 eIF4E2、eIF(iso)4E 和 nCBP 组成。在番茄中,eIF4E1 敲除 (KO) 可对一组马铃薯 Y 病毒组产生抗性,而 eIF4E1/2 双 KO 虽然可产生更广泛的抗性,但会导致植物发育缺陷。这里,四倍体马铃薯 cv。 Desirée 拥有显性 Ny 基因,该基因可抗马铃薯 Y 病毒 (PVY) 菌株 O 但不抗 NTN,用于评估通过 CRISPR-Cas9 介导的 eIF4E1 易感基因 KO 来扩大其 PVY 抗性谱的可能性。经过植物原生质体转染再生的双重过程,获得了 eIF4E1 KO 马铃薯。敲除是针对 eIF4E1 的,在其 eIF4E2 旁系同源物中未发现突变。eIF4E 家族的表达分析表明,eIF4E1 的破坏不会改变其他家族成员的 RNA 稳态水平。用 PVY NTN 分离物攻击的 eIF4E1 KO 系显示病毒积累减少和病毒诱导症状改善,表明 eIF4E1 基因是其增殖所必需的但不是必需的。我们的数据表明,可以通过增强 eIF4E 介导的隐性抗性,有效利用 eIF4E1 编辑来拓宽优良马铃薯品种(如 Desirée)的 PVY 抗性谱。
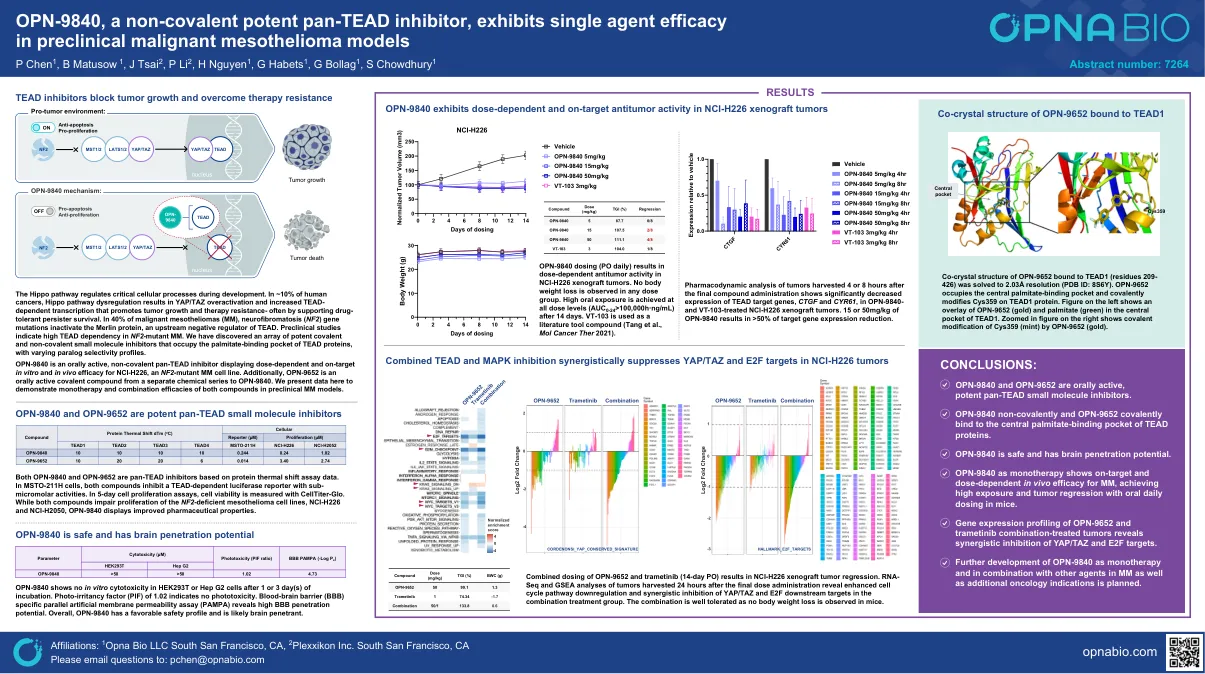
OPN-9840,一种非共价有效的泛胶抑制剂,在临床前恶性间皮瘤模型中表现出单药疗效
河马途径调节发育过程中关键细胞过程。在〜10%的人类癌症中,河马途径失调导致YAP/TAZ过度活化,并增加依赖性转录,从而促进肿瘤生长和耐药性 - 通常通过支持药物耐受性持久性生存。在40%的恶性间皮瘤(MM)中,神经纤维瘤病(NF2)基因突变使Merlin蛋白失活,Merlin蛋白是TEAD的上游负调节剂。临床前研究表明NF2突变MM中的高曲线依赖性。我们发现了一系列有效的共价和非共价小分子抑制剂,这些抑制剂占据了te蛋白的棕榈酸盐结合口袋,具有不同的旁系同源物选择性曲线。

HP1 是一种负责……的蛋白质,其新功能
真核生物的染色体由DNA和组蛋白组成,组蛋白的甲基化、乙酰化等化学修饰可诱导染色体聚集和松弛,从而改变基因表达模式。HP1已被证实为H3K9甲基化的结合蛋白,在促进染色体聚集中发挥作用。由于哺乳动物中HP1蛋白有3个旁系同源物,我们利用基因组编辑技术建立了3个HP1均缺失的细胞,并与正常细胞进行比较,发现在HP1缺陷细胞中,H3K9甲基转移酶和去甲基化酶大幅降解,染色体不能恢复成正确的结构(图1)。对部分功能缺失的HP1突变体的分析表明,HP1将H3K9甲基转移酶和去甲基化酶束缚在染色质上,阻止这些酶降解(图2)。

fpls-14-1247680.pdf - Rothamsted 存储库下载
简介:多酚氧化酶 (PPO) 是一种双活性金属酶,可催化醌的产生。在植物中,PPO 活性可能有助于抗生物胁迫和次生代谢,但对食品生产商来说是不利的,因为它会导致产品在收获后加工过程中变色和风味特征发生变化。在小麦 (Triticum aestivum L.) 中,在碾磨过程中从谷物的糊粉层释放出的 PPO 会导致面粉、面团和最终产品变色,从而降低其价值。同源组 2 染色体上的 PPO1 和 PPO2 旁系同源基因的功能丧失突变导致小麦粒中的 PPO 活性降低。然而,有限的自然变异和这些基因的接近性使得通过重组选择极低 PPO 小麦品种变得复杂。本研究的目标是编辑 PPO1 和 PPO2 的所有副本,以大幅降低优良小麦品种中的 PPO 籽粒活性。