XiaoMi-AI文件搜索系统
World File Search System甲状腺

研究人员更接近开发基于细胞的甲状腺功能减退疗法
“诱导的多能干细胞为再生医学提供了强大的工具,因为它们提供了一种无限的患者特异性细胞来源,可以潜在地用于治疗各种疾病。“这项研究使我们更接近使用IPSC衍生的甲状腺细胞进行甲状腺功能减退症的基于细胞的疗法。”

双相情感障碍与甲状腺功能障碍之间的相互作用:评论文章
双相情感障碍和甲状腺功能障碍通常表现出复杂的相互作用,会影响患者的临床表现和治疗结果。研究表明,甲状腺激素在情绪调节中起着至关重要的作用,甲状腺功能的波动会加剧或模仿双相情感障碍的症状。例如,甲状腺功能亢进症(其特征是甲状腺激素水平升高)可导致躁狂或轻躁狂发作,而甲状腺功能减退症(其激素分泌减少)可导致抑郁症状。这种错综复杂的关系需要对被诊断为双相情感障碍的患者的甲状腺功能进行全面评估,以解决任何可能恶化情绪稳定性的潜在激素失衡问题。此外,双相情感障碍的治疗可能需要谨慎管理甲状腺疾病,因为某些情绪稳定剂和抗抑郁药会影响甲状腺激素水平。据记录,通常用于治疗双相情感障碍的锂等药物既会影响甲状腺功能,又需要持续监测甲状腺相关的副作用。因此,医疗保健提供者应在整个治疗过程中对患有躁郁症的患者进行彻底的甲状腺功能评估。有效的管理策略应包括一种协作方法,包括精神科和内分泌科评估,以优化整体心理和身体健康。

甲状腺功能障碍,糖尿病和肥胖症患者的选定肌肉骨骼疾病
许多医疗状况会影响骨骼系统,并构成骨折的独立危险因素。甲状腺激素的作用对于维持足够的发育,矿化和骨骼强度是必要的。未经治疗的甲状腺功能亢进可导致骨矿物质密度(BMD),骨疏松症和病理骨折的降低。在甲状腺功能减退症中,骨骼结构质量的变化导致断裂频率的增加。 体重过多会对骨折的风险产生负面影响,增加骨关节炎的风险,并加速类风湿关节炎和骨质疏松症的发展。 1型和2型糖尿病与骨骼骨折的风险增加有关,尽管由于疾病的持续时间和促炎性状态,骨基质的持续时间不同,骨基质中晚期糖基化终产物(AGE)的局限性以及微血管疾病的疾病。 这项研究总结了有关甲状腺功能障碍,肥胖和糖尿病对骨骼系统的影响的当前文献。在甲状腺功能减退症中,骨骼结构质量的变化导致断裂频率的增加。体重过多会对骨折的风险产生负面影响,增加骨关节炎的风险,并加速类风湿关节炎和骨质疏松症的发展。1型和2型糖尿病与骨骼骨折的风险增加有关,尽管由于疾病的持续时间和促炎性状态,骨基质的持续时间不同,骨基质中晚期糖基化终产物(AGE)的局限性以及微血管疾病的疾病。这项研究总结了有关甲状腺功能障碍,肥胖和糖尿病对骨骼系统的影响的当前文献。

对孕产妇甲状腺功能减退症对胎儿发育的影响的叙述性回顾
甲状腺是位于人体颈部区域的蝴蝶形腺体。甲状腺产生三种激素,这些激素对于调节体温,能量产生,体重,头发和指甲生长以及月经周期维持至关重要。这些激素的产生由反馈机制控制。各种因素会导致这些激素的刺激和抑制变化,这最终会导致过度释放或甲状腺激素水平降低。这些原因可能是生理或病理。生理原因之一是怀孕。怀孕是一个非常复杂的过程,其中发生了许多变化及其功能。其中之一是母体甲状腺的变化。无法充分适应变化会导致甲状腺的异常功能。在怀孕期间,甲状腺激素的浓度有所不同,这可能导致甲状腺激素产生的水平或抑制作用降低。这种疾病称为甲状腺功能减退症。怀孕母亲的甲状腺功能减退症可以是妊娠,也可能是她怀孕前的情况。通常,妊娠甲状腺功能减退症在产后分娩后恢复,但也可以作为亚临床甲状腺功能减退症。在这种情况下,它们对发展构成了重大威胁,对子宫中的婴儿造成了障碍,并在将来导致后代异常。因此,母体激素水平的干扰干扰了典型的早期发育特征。由于甲状腺结合蛋白水平的增强,肾脏中碘的清除率提高,肾脏中碘的清除率提高,人类绒毛膜促性腺激素激素的影响改变,以及饮食中碘的消耗降低,发生了一些变化。 孕产妇甲状腺功能减退症中的碘失衡与严重的健康问题有关,例如果仁糖浆和智力低下。 甲状腺激素对子宫中婴儿的神经,认知和智力商发育至关重要。 在迅速发展的科学研究的世界中,可以通过多种方式可以早日检测到这种情况,并得到正确的诊断,并给予恰当的和必要的关注和治疗,从而对胎儿和母亲造成最小的伤害。发生了一些变化。孕产妇甲状腺功能减退症中的碘失衡与严重的健康问题有关,例如果仁糖浆和智力低下。 甲状腺激素对子宫中婴儿的神经,认知和智力商发育至关重要。 在迅速发展的科学研究的世界中,可以通过多种方式可以早日检测到这种情况,并得到正确的诊断,并给予恰当的和必要的关注和治疗,从而对胎儿和母亲造成最小的伤害。孕产妇甲状腺功能减退症中的碘失衡与严重的健康问题有关,例如果仁糖浆和智力低下。甲状腺激素对子宫中婴儿的神经,认知和智力商发育至关重要。在迅速发展的科学研究的世界中,可以通过多种方式可以早日检测到这种情况,并得到正确的诊断,并给予恰当的和必要的关注和治疗,从而对胎儿和母亲造成最小的伤害。

孔五角形 - 甲状腺石墨烯:锂离子电池的有前途的阳极材料
作者的完整列表:杨,布伦达;约翰·霍普金斯医学鲁特科夫斯基,娜塔莉; Johns Hopkins Medicine Elisseeff,Jennifer;约翰·霍普金斯大学,生物医学工程

甲状腺营养ANME-1古细菌的进化多样化及其膨胀病毒蛋白
5 Albers,S. -V。 &Jarrell,K。F.古细菌:古细菌如何游泳。 微生物学中的边界6,doi:10.3389/fmicb.2015.00023(2015)。 6 Albers,S. -V。 &Jarrell,K。F. Archaellum:独特的古细菌运动结构的更新。 微生物学的趋势26,351-362,doi:https://doi.org/10.1016/j.tim.2018.01.004(2018)。 7 Van Wolferen,M.,Orell,A。 &Albers,S. -V。 古细菌生物膜形成。 自然评论微生物学16,699-713(2018)。 8 Pohlschroder,M。&Esquivel,R。N.古细菌IV pili及其参与生物膜形成。 微生物学的前沿6,190(2015)。 9 Walker,D。等。 hungatei的甲螺旋藻的古细胞是导电性的。 。 MBIO 10,E00579-00519(2019)。 10 Holmes,D。E.,Zhou,J.,Ueki,T.,Woodard,T。&Lovley,D。R.在直接种间电子传输过程中,甲那粒乙酸盐乙酸盐摄取电子的机制。 MBIO 12,E02344-02321(2021)。 11 Quemin,E。R.等。 首先深入了解过度授予性古细菌病毒的进入过程。 J Virol 87,13379-13385,doi:10.1128/jvi.02742-13(2013)。 12 Baquero,D。P.等。 病毒研究的进展。 108(eds Margaret Kielian,Thomas C. Mettenleiter和Marilyn J. Roossinck)127-164(学术出版社,2020年)。 13 Briegel,A。等。 跨古细菌和细菌的趋化机制的结构保护。 环境微生物学报告7,414-419,doi:https://doi.org/10.1111/1758-222299.12265(2015)。5 Albers,S. -V。&Jarrell,K。F.古细菌:古细菌如何游泳。微生物学中的边界6,doi:10.3389/fmicb.2015.00023(2015)。6 Albers,S. -V。 &Jarrell,K。F. Archaellum:独特的古细菌运动结构的更新。 微生物学的趋势26,351-362,doi:https://doi.org/10.1016/j.tim.2018.01.004(2018)。 7 Van Wolferen,M.,Orell,A。 &Albers,S. -V。 古细菌生物膜形成。 自然评论微生物学16,699-713(2018)。 8 Pohlschroder,M。&Esquivel,R。N.古细菌IV pili及其参与生物膜形成。 微生物学的前沿6,190(2015)。 9 Walker,D。等。 hungatei的甲螺旋藻的古细胞是导电性的。 。 MBIO 10,E00579-00519(2019)。 10 Holmes,D。E.,Zhou,J.,Ueki,T.,Woodard,T。&Lovley,D。R.在直接种间电子传输过程中,甲那粒乙酸盐乙酸盐摄取电子的机制。 MBIO 12,E02344-02321(2021)。 11 Quemin,E。R.等。 首先深入了解过度授予性古细菌病毒的进入过程。 J Virol 87,13379-13385,doi:10.1128/jvi.02742-13(2013)。 12 Baquero,D。P.等。 病毒研究的进展。 108(eds Margaret Kielian,Thomas C. Mettenleiter和Marilyn J. Roossinck)127-164(学术出版社,2020年)。 13 Briegel,A。等。 跨古细菌和细菌的趋化机制的结构保护。 环境微生物学报告7,414-419,doi:https://doi.org/10.1111/1758-222299.12265(2015)。6 Albers,S. -V。&Jarrell,K。F. Archaellum:独特的古细菌运动结构的更新。微生物学的趋势26,351-362,doi:https://doi.org/10.1016/j.tim.2018.01.004(2018)。7 Van Wolferen,M.,Orell,A。&Albers,S. -V。古细菌生物膜形成。自然评论微生物学16,699-713(2018)。8 Pohlschroder,M。&Esquivel,R。N.古细菌IV pili及其参与生物膜形成。微生物学的前沿6,190(2015)。9 Walker,D。等。hungatei的甲螺旋藻的古细胞是导电性的。。MBIO 10,E00579-00519(2019)。 10 Holmes,D。E.,Zhou,J.,Ueki,T.,Woodard,T。&Lovley,D。R.在直接种间电子传输过程中,甲那粒乙酸盐乙酸盐摄取电子的机制。 MBIO 12,E02344-02321(2021)。 11 Quemin,E。R.等。 首先深入了解过度授予性古细菌病毒的进入过程。 J Virol 87,13379-13385,doi:10.1128/jvi.02742-13(2013)。 12 Baquero,D。P.等。 病毒研究的进展。 108(eds Margaret Kielian,Thomas C. Mettenleiter和Marilyn J. Roossinck)127-164(学术出版社,2020年)。 13 Briegel,A。等。 跨古细菌和细菌的趋化机制的结构保护。 环境微生物学报告7,414-419,doi:https://doi.org/10.1111/1758-222299.12265(2015)。MBIO 10,E00579-00519(2019)。10 Holmes,D。E.,Zhou,J.,Ueki,T.,Woodard,T。&Lovley,D。R.在直接种间电子传输过程中,甲那粒乙酸盐乙酸盐摄取电子的机制。MBIO 12,E02344-02321(2021)。11 Quemin,E。R.等。首先深入了解过度授予性古细菌病毒的进入过程。J Virol 87,13379-13385,doi:10.1128/jvi.02742-13(2013)。12 Baquero,D。P.等。病毒研究的进展。108(eds Margaret Kielian,Thomas C. Mettenleiter和Marilyn J. Roossinck)127-164(学术出版社,2020年)。13 Briegel,A。等。跨古细菌和细菌的趋化机制的结构保护。环境微生物学报告7,414-419,doi:https://doi.org/10.1111/1758-222299.12265(2015)。14 Quax,T。E. F.,Albers,S. -V。 &Pfeiffer,古细菌的出租车。 生命科学的新兴主题2,535-546,doi:10.1042/etls20180089(2018)。 15 Li,Z.,Rodriguez -Franco,M.,Albers,S. -V。 &Quax,T。E. F.开关复合物Arlcde连接趋化系统和古细胞。 分子微生物学114,468-479,doi:https://doi.org/10.1111/mmi.14527(2020)。 16 Meyerdierks,A。等。 元基因组和mRNA表达分析ANME -1组的厌氧性古细菌。 环境微生物学12,422-439,doi:doi:10.1111/j.1462-2920.2009.02083.x(2010)。 17 Chadwick,G。L.等。 比较基因组学揭示了电子转移和综合机制,从而区分了甲状腺营养和甲烷古细菌。 PLOS生物学20,E3001508,doi:10.1371/journal.pbio.3001508(2022)。 18 Zheng,K.,Ngo,P。D.,Owens,V。L.,Yang,X. -P。 &Mansoorabadi,S。O。 甲酶F430在甲烷和甲状腺营养古细菌中的生物合成途径。 Science 354,339-342,doi:10.1126/science.aag2947(2016)。 19 Michael,A。J.多胺在古细菌和细菌中的功能。 生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。 20 Morimoto,N。等。 在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。 细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。 21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。14 Quax,T。E. F.,Albers,S. -V。&Pfeiffer,古细菌的出租车。生命科学的新兴主题2,535-546,doi:10.1042/etls20180089(2018)。15 Li,Z.,Rodriguez -Franco,M.,Albers,S. -V。 &Quax,T。E. F.开关复合物Arlcde连接趋化系统和古细胞。 分子微生物学114,468-479,doi:https://doi.org/10.1111/mmi.14527(2020)。 16 Meyerdierks,A。等。 元基因组和mRNA表达分析ANME -1组的厌氧性古细菌。 环境微生物学12,422-439,doi:doi:10.1111/j.1462-2920.2009.02083.x(2010)。 17 Chadwick,G。L.等。 比较基因组学揭示了电子转移和综合机制,从而区分了甲状腺营养和甲烷古细菌。 PLOS生物学20,E3001508,doi:10.1371/journal.pbio.3001508(2022)。 18 Zheng,K.,Ngo,P。D.,Owens,V。L.,Yang,X. -P。 &Mansoorabadi,S。O。 甲酶F430在甲烷和甲状腺营养古细菌中的生物合成途径。 Science 354,339-342,doi:10.1126/science.aag2947(2016)。 19 Michael,A。J.多胺在古细菌和细菌中的功能。 生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。 20 Morimoto,N。等。 在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。 细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。 21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。15 Li,Z.,Rodriguez -Franco,M.,Albers,S. -V。&Quax,T。E. F.开关复合物Arlcde连接趋化系统和古细胞。分子微生物学114,468-479,doi:https://doi.org/10.1111/mmi.14527(2020)。16 Meyerdierks,A。等。元基因组和mRNA表达分析ANME -1组的厌氧性古细菌。环境微生物学12,422-439,doi:doi:10.1111/j.1462-2920.2009.02083.x(2010)。17 Chadwick,G。L.等。 比较基因组学揭示了电子转移和综合机制,从而区分了甲状腺营养和甲烷古细菌。 PLOS生物学20,E3001508,doi:10.1371/journal.pbio.3001508(2022)。 18 Zheng,K.,Ngo,P。D.,Owens,V。L.,Yang,X. -P。 &Mansoorabadi,S。O。 甲酶F430在甲烷和甲状腺营养古细菌中的生物合成途径。 Science 354,339-342,doi:10.1126/science.aag2947(2016)。 19 Michael,A。J.多胺在古细菌和细菌中的功能。 生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。 20 Morimoto,N。等。 在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。 细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。 21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。17 Chadwick,G。L.等。比较基因组学揭示了电子转移和综合机制,从而区分了甲状腺营养和甲烷古细菌。PLOS生物学20,E3001508,doi:10.1371/journal.pbio.3001508(2022)。18 Zheng,K.,Ngo,P。D.,Owens,V。L.,Yang,X. -P。 &Mansoorabadi,S。O。 甲酶F430在甲烷和甲状腺营养古细菌中的生物合成途径。 Science 354,339-342,doi:10.1126/science.aag2947(2016)。 19 Michael,A。J.多胺在古细菌和细菌中的功能。 生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。 20 Morimoto,N。等。 在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。 细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。 21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。18 Zheng,K.,Ngo,P。D.,Owens,V。L.,Yang,X. -P。&Mansoorabadi,S。O。甲酶F430在甲烷和甲状腺营养古细菌中的生物合成途径。Science 354,339-342,doi:10.1126/science.aag2947(2016)。19 Michael,A。J.多胺在古细菌和细菌中的功能。 生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。 20 Morimoto,N。等。 在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。 细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。 21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。19 Michael,A。J.多胺在古细菌和细菌中的功能。生物学杂志293,18693-18701,doi:https://doi.org/10.1074/jbc.tm118.005670(2018)。20 Morimoto,N。等。在高疗法中的长链多胺的双重生物合成途径 thermoccus kodakarensis 。细菌学杂志192,4991-5001,doi:doi:10.1128/jb.00279-10(2010)。21 Kanehisa,M。&Goto,S。Kegg:基因和基因组的京都百科全书。核酸研究28,27-30,doi:10.1093/nar/28.1.27(2000)。22 Mihara,H。&Esaki,N。细菌半胱氨酸脱硫酶:它们的功能和机制。应用微生物学和生物技术60,12-23,doi:10.1007/s00253-002-1107-4(2002)。23 Tchong,S.-I.,Xu,H。&White,R。H. L-半胱氨酸脱硫酶:一种从Jannaschii中分离出的[4FE -4S]酶,催化了L-半结合体为吡酸丙酮酸,氨氨基和硫化物的溶解。生物化学44,1659-1670,doi:10.1021/bi0484769(2005)。

国家临床卓越研究所
经常发生在Graves疾病(一种自身免疫性疾病)的同时,导致免疫系统过度刺激甲状腺,从而导致甲状腺功能亢进。大约40%的坟墓疾病患者受到TED的影响。1在坟墓疾病中,称为甲状腺刺激免疫球蛋白(TSIS)的抗体不仅刺激甲状腺产生过多的甲状腺激素,而且还可以攻击眼睛后面的组织,从而导致TED。TED的独特特征之一是它独立于甲状腺功能的能力。虽然在患有坟墓疾病和甲状腺功能亢进症的个体中看到许多TED病例,但TED也可以表现出在甲状腺功能正常(甲状腺功能纤维)甚至甲状腺功能不足(甲状腺功能减退)的患者中。在全球范围内,TED患者的甲状腺功能亢进,甲状腺功能减退症和甲状腺功能亢进的患病率分别为86.2%,10.36%和7.9%。2
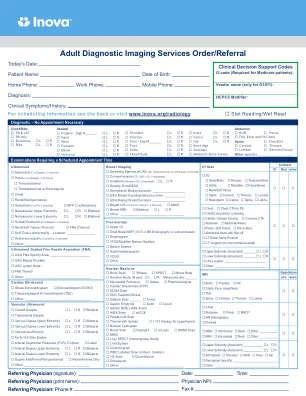
成人诊断成像服务订单/推荐
核医学骨骼扫描:SpectImpost:乳房:乳房:themant:ther:LR黑色素瘤部位:_________________________________药理学药理药理学心疗中心病(pyp)(pyp)固体液体胃反流(牛奶扫描)HIDA扫描w/cck甲状旁腺扫描甲状腺甲状腺甲状腺甲状腺甲状腺甲状腺甲状腺甲状腺甲状腺甲状腺疗法核囊图:肾脏囊力术:肾脏扫描:肾脏扫描:in Liver/Spleen Cisternogram WBC Labeled Scan (Indium, Ceretec) VQ Scan Quantitative Octreoscan Other: _____________________________________

2025年5月3日和4日
甲状腺机器人手术功能性甲状腺疾病的甲状腺眼疾病新颖的疗法,可在良性和恶性甲状腺疾病中进行手术进展和微创手术消融疗法,以评估甲状腺结节的人性化疾病,并评估甲状腺疾病的疾病,并评估甲状腺疾病的先进癌症,并评估成人癌症的疾病,并评估甲状腺疾病的先进癌症,并用于治疗MTC酪氨酸激酶抑制剂和靶向治疗的基因分析,用于甲状腺功能亢进症的甲状腺癌手术治疗ATA 2025年甲状腺结节和癌症的指南

放射治疗
接受过部分或全部甲状腺切除术或甲状腺消融术的患者将接受左旋甲状腺素替代治疗,以避免甲状腺功能减退,并将促甲状腺激素 (TSH) 水平保持在正常范围内。由于甲状腺组织和甲状腺癌细胞需要高水平的 TSH 才能吸收放射性碘 I-131,因此患者可以在扫描/治疗前 2 周服用左旋甲状腺素,或接受重组形式的 TSH(促甲状腺激素 α)。这是肌肉注射的 - 每天一次,持续两天(参见 HNOTTSH 方案)。如果使用促甲状腺激素 α,患者不必服用左旋甲状腺素,也不必冒着出现甲状腺功能减退症状的风险。有关血清甲状腺球蛋白 (Tg) 和 TSH 测试的信息可以在 BC 癌症药物手册的促甲状腺激素 α 专论中找到。