XiaoMi-AI文件搜索系统
World File Search System直系

1 电子通信法内容...
(3) 委员会成员就职时,应提交关于存在或不存在重大商业、金融或其他商业利益的声明。声明还应包含与委员会成员或其家庭成员有经济关系的人员的姓名和地址,无限制的直系血亲,四代以内的旁系亲属,二代以内的姻亲,以及由此产生的商业利益。声明应保存在委员会的特别公共登记册中

GENESPACE 追踪多个基因组中的感兴趣区域和基因拷贝数变异
摘要 许多分类群中多个染色体规模的参考基因组序列的开发已产生对分子进化模式和过程的高分辨率视图。尽管如此,利用跨多个基因组的信息仍然是几乎所有真核生物系统中的重大挑战。这些挑战包括研究染色体结构的进化、寻找数量性状基因座的候选基因以及检验有关物种形成和适应的假设。在这里,我们提出了 GENESPACE,它通过整合保守的基因顺序和直系同源性来解决这些挑战,以确定所有基因在多个基因组中的预期物理位置。我们通过从三个生物组织水平剖析存在-缺失、拷贝数和结构变异来证明这一实用性:跨越 3 亿年的脊椎动物性染色体进化、跨禾本科(草类)植物家族的多样性以及 26 个玉米品种。 GENESPACE R 包中构建和可视化同源直系同源性的方法为现有的基因家族和同源性程序提供了重要的补充,特别是在多倍体、杂交和其他复杂基因组中。
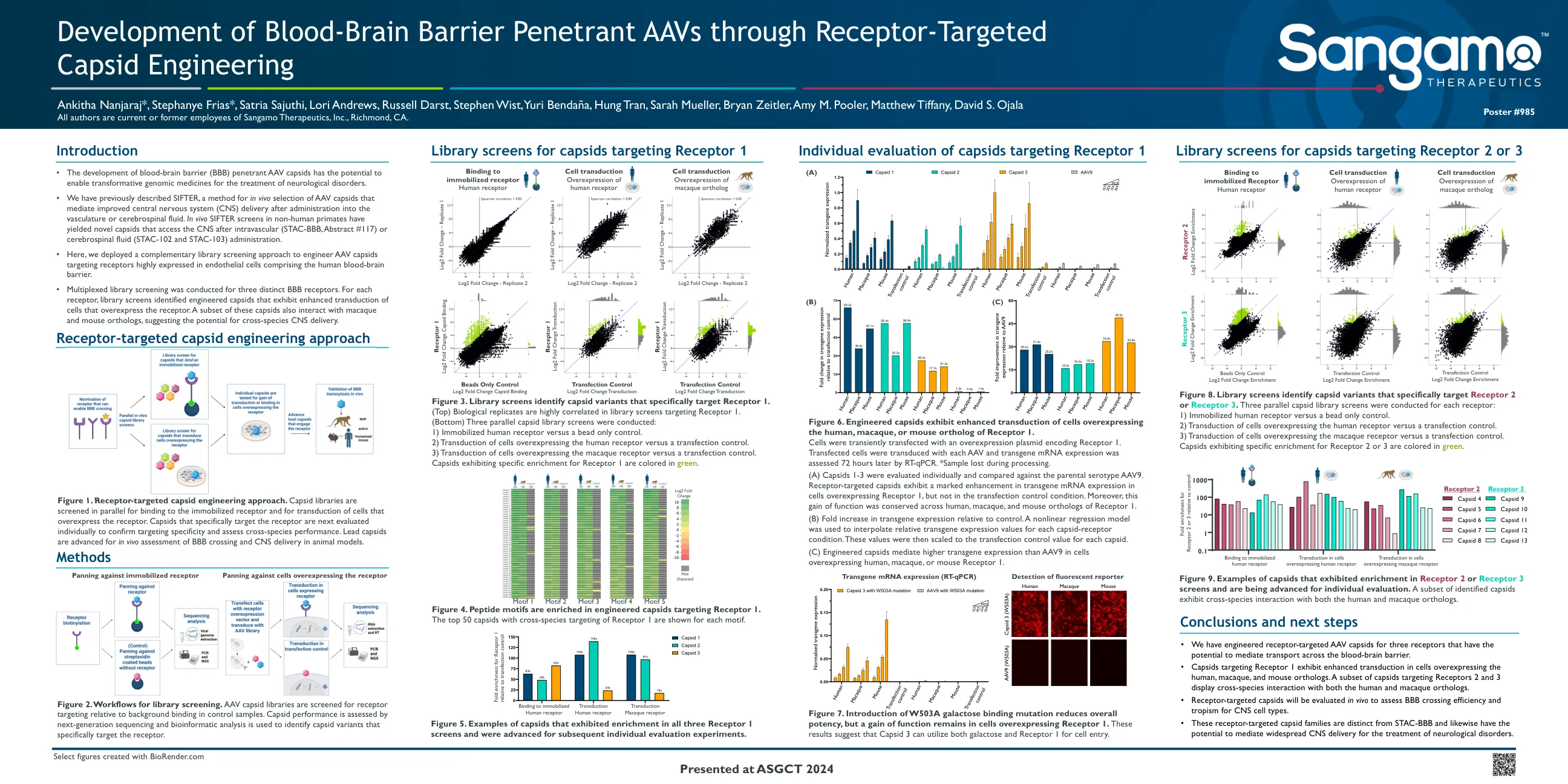
靶向受体的衣壳工程方法...
图6。工程的衣壳表现出增强的过表达人,猕猴或小鼠直系同源物的细胞的转导增强。细胞用过表达质粒编码受体1。转染的细胞用每种AAV转导,并在72小时后通过RT-QPCR评估转基因mRNA表达。*处理过程中丢失的样本。(a)CAPSIDS 1-3单独评估并与父母血清型AAV9进行比较。受体靶向的衣壳在过表达受体1的细胞中表现出明显的转基因mRNA表达,但在转染控制条件下却没有明显增强。此外,这种功能的增益在受体的人,猕猴和小鼠直系同源物中得到了保守。(b)相对于对照的转基因表达的倍数增加。一个非线性回归模型用于每个衣壳受体条件的相对转基因表达值。然后将这些值缩放到每个衣壳的转染控制值。(C)在过表达人,猕猴或小鼠受体1的细胞中,工程上的衣壳介导更高的转基因表达。

数据发布 利用染色体长度的基因组组装来注释亚洲柑橘木虱(Diaphorina citri)的 Wnt 信号通路
表 2. 支持基因注释的证据。手工注释的柑橘木虱 Wnt 通路基因。总共有 24 个基因模型。每个基因模型都分配了一个标识符,并列出了用于验证或修改基因模型结构的证据。还列出了最能支持手工注释的 MCOT 转录组标识符。当存在从头转录组、Iso-Seq、RNA-Seq 和直系同源物支持的证据时,表中会标记“X”。MCOT:基于基因组 MAKER、Cufflinks、Oases 和 Trinity 转录本预测的综合转录组;MAKER:基因预测;从头转录组:使用 Iso-Seq 长读和 RNA-Seq 数据的独立转录组;Iso-Seq 转录本:用 Pacific Biosciences 技术生成的全长转录本; RNA-Seq:映射到基因组的读取也用作剪接点的支持证据;直系同源物证据:来自相关半翅目物种和果蝇的蛋白质。

用于神经退行性实验模型的人工智能
图 1 人类与非人类物种之间共享的基因。系统发育树标注了每个物种中具有 1:1 直系同源物的人类基因百分比(以数字和每个圆圈的填充比例显示)。与人类共享的 1:1 直系同源物的绝对数量绘制为每个圆圈的颜色。使用 orthogene R 包构建。92 关键词:Anolis carolinensis,绿变色蜥;Bos taurus,牛;Caenorhabditis elegans,蛔虫;Canis lupus familiaris,狗;Danio rerio,斑马鱼;Drosophila melanogaster,果蝇;Equus caballus,马;Felis catus,猫;Gallus gallus,鸡;Homo sapiens,人类;Macaca mulatta,恒河猴;Monodelphis domestica,灰色短尾负鼠;小家鼠 (Mus musculus),家鼠;鸭嘴兽 (Ornithorhynchus anatinus),鸭嘴兽;黑猩猩 (Pan troglodytes),黑猩猩;褐家鼠 (Rattus norvegicus),褐家鼠;酿酒酵母 (Saccharomyces cerevisiae),面包酵母;粟酒裂殖酵母 (Schizosaccharomyces pombe),裂殖酵母;野猪 (Sus scrofa),猪;热带爪蟾 (Xenopustropicalis),西方爪蟾。

植物抗癌药物的代谢工程
测序和转录组学的进步使得通过共表达分析可以发现酶,其中候选基因通过组织表达模式与已知途径酶的相似性来识别 — — 最近在 C.roseus 和 Podophyllumpeltatum 中的发现证明了这一点 [ 4 , 5 ]。自组织映射等机器学习方法进一步优化了候选基因 [ 6 ]。这些方法,加上对植物体内生物合成定位的更深入理解,以及单细胞代谢组学等技术的发展,进一步改善了候选基因的选择,加速了酶的发现 [ 7 ]。借助基于 OMIC 的工具(如 plantiSMASH)识别物理基因簇有助于阐明缺失的生物合成酶,如那可丁和长春花碱途径中的酶 [ 8–10 ]。然而,这种方法是有限的,因为许多植物生物合成途径几乎没有或没有基因聚集,如喜树碱生物合成途径[11]。基于同源性的克隆可以加速发现与已知生物合成酶具有直系同源功能的基因,例如在 Tabernanthe iboga 的 ibogaine 生物合成途径中鉴定出 C. roseus 脱羧酶直系同源物[12]。然而,途径的复杂性往往需要采用组合方法,例如 Gelsemiumsempervirens 氧化吲哚途径的发现[13]。

使用疟原虫作为筛选疟原虫血液阶段疟疾疫苗靶标的模型揭示了新的候选
疟原虫造成非洲以外的大多数疟疾病例。与p不同。恶意,p。Vivax生命周期包括休眠的肝脏,催眠症,在没有蚊子传播的情况下会引起感染。一种针对p的有效疫苗。Vivax血液阶段将限制这种复发感染的症状和病理,因此可能在控制该物种的控制中起关键作用。p。vivax落后于p。恶性菌,有许多识别的tar-得到了几个转换为II期测试。相比之下,只有一个p。基于Divax血液阶段疫苗基于DUFFY结合蛋白(PVDBP)的候选疫苗已达到IA期,这在很大程度上是因为缺乏p的持续体外培养系统。Vivax限制了新候选人的系统筛选。我们使用了p之间的密切系统发育关系。vivax和p。knowlesi(人类红细胞中存在体外培养系统),以测试系统反疫苗学的可扩展性以识别和确定p的优先级。Vivax血液阶段目标。p。在哺乳动物的表达系统中,预测在红细胞侵袭中起作用的可在红细胞侵袭中起作用。 这些抗原中的八种用于产生多克隆抗体,这些抗体被筛选,以识别p中的直系同源蛋白的能力。 knowlesi。 knowlesi和嵌合p。 knowlesi基因与他们的p。在哺乳动物的表达系统中,预测在红细胞侵袭中起作用的可在红细胞侵袭中起作用。这些抗原中的八种用于产生多克隆抗体,这些抗体被筛选,以识别p中的直系同源蛋白的能力。knowlesi。knowlesi和嵌合p。knowlesi基因与他们的p。然后对这些抗体进行了测试,以抑制两种野生型P的生长和侵袭。使用CRISPR/CAS9进行修改以交换p。Vivax直系同源物。诱导抑制抗体的候选者

解决人类和黑猩猩神经发育过程中人类加速区域的三维相互作用
抽象的人类加速区域(HAR)在物种之间是高度保守的,但表现出显着的人类特异性序列变化,这表明它们可能在人类进化中获得了新的功能。hars包括具有人类特异性活性的转录增强子,并与人脑的进化有关。然而,我们对Hars如何促进大脑独特的人类特征的理解受到了对Har har统治的基因和途径的洞察力的阻碍。目前尚不清楚Hars是通过改变har及其黑猩猩直系同源物之间的基因靶标的表达而作用的,还是通过在人类中获得新的基因靶标,这是一种称为增强子劫持的机制。,我们在人和黑猩猩神经干细胞(NSC)中生成了1,590个har及其直系同源物的染色质相互作用的高分辨率图,以全面地识别这两个物种中的基因靶标。hars及其嵌合式直系同源物的目标是一组2,963个基因,这些基因富含神经发育过程,包括神经发生和突触传播。HAR增强剂活性的变化与保守基因靶标表达的变化相关。保守的靶标在人与黑猩猩NSC之间或人类和非人类灵长类动物发展和成人大脑之间差异表达的基因中富集。特异性的HAR基因靶标未在已知的生物学功能上融合,并且在差异表达的基因中没有显着富集,这表明Hars不会通过增强子劫持来改变基因表达。har Gene靶标,包括差异表达的靶标,还显示了发育中的人脑中的细胞类型特异性表达模式,包括外部径向胶质细胞,这些模式有助于人类皮质膨胀。我们的发现支持Hars通过改变保守基因靶标的表达来影响人脑的演变,并提供与新型人类大脑特征联系起来的手段。

CRISPR介导的转录抑制作用弓形虫Gondii
用于调整内源基因表达的抽象工具是确定各种细胞表型的遗传基础的关键。尽管在弓形虫中可以使用合成的可调节性,但基因表达的靶向和组合下调的可扩展方法(如RNA干扰)尚未得到发展。为了研究CRISPR介导的转录调控的可行性,我们研究了来自甲状腺链球菌和嗜热链球菌的两个催化无活性Cas9(DCAS9)直系同源物的功能。在添加靶向启动子的单个指定RNA(SGRNA)和表面抗原基因SAG1的5 9个未翻译区域(UTR)后,我们对靶基因的蛋白质刺激蛋白质的变化通过流量细胞仪的蛋白质刺激变化,用于转录报告基因和immu-immu-nosu-nosu-noce。我们发现DCAS9直系同源物产生了一系列靶基因表达水平,并且抑制程度持久且稳定地遗传。因此,化脓性链球菌和嗜热链球菌DCAS9可以有效地产生弓形虫中的基因表达水平。两个DCAS9的独特的SGRNA支架要求允许通过转录调制,基于显微镜的研究标记或其他基于DCAS9的方法同时检查两个不同的基因座。利用新近获得的基因组转录起始站点数据,这些工具将有助于开发弓形虫的新功能筛查方法。
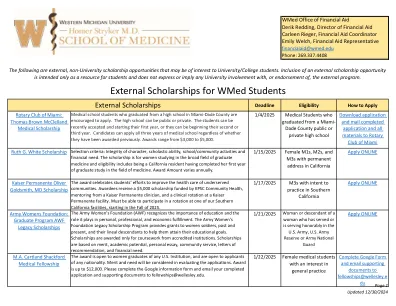
WMED学生的外部奖学金
陆军妇女基金会(AWF)认识到教育的重要性及其在个人,专业和经济实现中所扮演的角色。陆军妇女基金会遗产奖学金计划为过去和现在的女士兵及其直系后代提供赠款,以帮助她们实现自己的教育目标。奖学金仅因获得认可机构的课程而授予奖学金。奖学金是基于功绩,学术潜力,个人论文,社区服务,推荐信和财务需求。