XiaoMi-AI文件搜索系统
World File Search System编辑器

碱基编辑器:在活细胞中引入点突变的模块化工具
碱基编辑器是一类新的可编程基因组编辑工具,它将 ssDNA(单链 DNA)修饰酶与催化失活的 CRISPR 相关(Cas)核酸内切酶融合,以诱导高效的单碱基变化。目前已报道了数十种碱基编辑器,显然这些工具是高度模块化的;ssDNA 修饰酶和 Cas 蛋白的多种组合产生了各种碱基编辑器,每种编辑器都有其独特的属性和潜在用途。从这个角度来看,我们描述了当前可用的碱基编辑器,强调了它们的模块化特性并描述了每个组件可用的各种选项。此外,我们简要讨论了合成生物学和基因组工程中的应用,在这些应用中,碱基编辑器比其他技术具有独特的优势。

植物引物编辑器实现水稻细胞的精准基因编辑
基因组编辑正在彻底改变植物研究和作物育种。序列特异性核酸酶 (SSN),例如锌指核酸酶 (ZFN) 和 TAL 效应核酸酶 (TALEN),已用于产生位点特异性 DNA 双链断裂并通过促进同源定向修复 (HDR) 实现精确的 DNA 修饰 (Steinert 等人,2016 年;Voytas,2013 年)。后来,RNA 引导的 SSN,例如 CRISPR-Cas9、Cas12a、Cas12b 及其变体,已应用于植物基因组编辑 (Li 等人,2013 年;Nekrasov 等人,2013 年;Tang 等人,2017 年;Zhong 等人,2019 年;Ming 等人,2020 年;Tang 等人,2019 年)。然而,HDR 依赖于 SSN 和 DNA 供体的同时递送,这在植物中一直具有挑战性( Steinert 等,2016; Zhang 等,2019)。在植物中实现高效 HDR 的另一个挑战是,在大多数细胞类型中,DNA 修复倾向于非同源末端连接(NHEJ)途径而不是 HDR( Puchta,2005; Qi 等,2013)。与受供体选择和 DNA 修复机制限制的 SSN 诱导的 HDR 不同,近年来开发的胞苷或腺嘌呤碱基编辑器可以在原型间隔物中 3-8 个核苷酸靶向窗口内将 C 转换为 T 或将 A 转换为 G( Komor 等,2016; Nishida 等,2016; Gaudelli 等,2017)。碱基编辑器虽然效率很高,但只能指导某些转换突变,而不能执行预定的颠换突变或插入和缺失 (indel)。在所有这些背景下,最近在人类细胞中开发所谓的引物编辑器 (PE) 方面取得的突破非常令人兴奋 ( Anzalone 等人,2019 )。在引物编辑中,Cas9H840A 切口酶与逆转录酶融合。融合蛋白在编辑 DNA 链上切口,通过引导到切口 DNA 并复制由引物编辑向导 RNA (pegRNA) 编码的遗传信息来启动逆转录。多功能的 pegRNA 是一种经过修饰的单向导 RNA (sgRNA),其 3' 端携带逆转录 (RT) 模板和引物结合位点 (PBS) 或序列中的引物。与 HDR 不同,PE 不需要 DNA 供体。在某些目标位点,PE 似乎也比碱基编辑器更精确、更高效(Anzalone 等人,2019 年)。

高保真碱基编辑器,没有可检测的全基因组脱靶效应
预印本(未经同行评审认证)是作者/资助者。保留所有权利。未经许可不得重复使用。此版本的版权所有者于 2020 年 2 月 9 日发布。;https://doi.org/10.1101/2020.02.07.939074 doi:bioRxiv 预印本
适用于 Spirent GNSS Constellation 的 SimGEN® 软件套件...
图 1 基于 SimGEN 的 GSS9000 GNSS 仿真系统示例 .............................................................................. 8 图 2 GSS7000 GNSS 仿真系统示例 .............................................................................................. 8 图 3 SimGEN 图形用户界面示例 ...................................................................................................... 9 图 4 场景树 ...................................................................................................................................... 10 图 5 车辆(天线)位置、运动和接收信号显示 ............................................................................. 11 图 6 卫星地面轨迹和天空图 ............................................................................................................. 11 图 7 典型的源编辑器 ................................................................................................................ 12 图 8 典型的星座编辑器 – 显示 GPS ............................................................................................. 13 图 9 卫星地面轨迹 ............................................................................................................................. 14 图 10 信号内容定义 – 显示 GPS ............................................................................................. 15 图 11 大气模型系数 ................................................................................................................ 16 图 12 定义车辆性能范围的个性编辑器 ................................................................................. 18 图13 赛道编辑器 ................................................................................................................................ 19 图 14 圆周运动编辑器 ...................................................................................................................... 20 图 15 飞机运动命令编辑器 ................................................................................................................ 21 图 16 航天器位置编辑器 ................................................................................................................ 24 图 17 地形遮挡编辑器 ...................................................................................................................... 26 图 18 天线模式编辑器 ...................................................................................................................... 27 图 19 天线杠杆臂 ............................................................................................................................. 27 图 20 Sim3D™ 环境表示 ................................................................................................................ 28 图 21 统计多径类别掩模编辑器 ............................................................................................................. 29 图 22 GTx 的功率与距离建模 ..................................................................................................... 31 图 23 快速查看选择和记录 ............................................................................................................. 32 图 24 数据流 ............................................................................................................................. 33 图 25 信号类型选择 ............................................................................................................................. 34 图 26 GBAS 消息类型 1 和 2 编辑器示例 ...................................................................................... 38
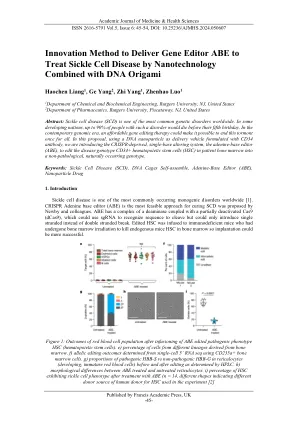
创新方法提供基因编辑器ABE来治疗纳米技术与DNA折纸结合创新方法提供基因编辑器ABE来治疗纳米技术与DNA折纸结合
治疗方法很好,但是治疗的输送方法在临床上并不可行。接受HSC治疗的受体小鼠必须接受致命剂量的γ辐射,以消除大量的骨髓(其中包含绝大多数造血干细胞,HSC),从而带来了重大的安全问题。可能更希望寻求方法,而无需破坏骨髓中HSC的原始菌落。一种方法是制定腺嘌呤碱基编辑器(ADE),并通过静脉内途径将其传递到骨髓中,以实现体内编辑。图1显示了有关治疗小鼠的一些统计数据,包括β球蛋白的百分比,细胞形态等。可以清楚地看出,经过治疗的小鼠的细胞已恢复正常的形态,证明了这种治疗的有效性。

高通量变体库和机器学习为 Retron 基因编辑器提供设计规则
细菌逆转录酶系统在许多生物技术应用中充当单链 DNA 的细胞内工厂。在这些技术中,天然的逆转录酶非编码 RNA (ncRNA) 被修饰以编码模板,以通过逆转录产生定制 DNA 序列。逆转录效率是逆转录酶技术的主要限制步骤,但我们缺乏系统的知识,了解如何在改变逆转录酶序列以产生定制 DNA 的同时提高或保持逆转录效率。在这里,我们测试了数千种对逆转录酶-Eco1 ncRNA 的不同修饰,并在汇集变体文库实验中测量 DNA 的产生,从而确定了 ncRNA 中对修饰具有耐受性和不耐受性的区域。我们将这些新信息应用于特定应用:使用逆转录酶与 CRISPR-Cas9 RNA 引导核酸酶 (editron) 结合产生精确的基因组编辑供体。我们使用酿酒酵母中的高通量文库来额外定义编辑酶的设计规则。我们将有关 retron DNA 生成和编辑子设计规则的新知识扩展到人类基因组编辑,以实现迄今为止最高效率的 retron-Eco1 编辑子。

Med surg made incredible easy pdf 第 8 版在线 pdf 编辑器
作为居住在德国的外籍人士,了解您的资金管理选项可能很棘手。从开设银行账户到为家人的房屋和财产投保,了解哪些选项适合您非常重要。要了解如何让您的资金发挥更大的作用,请阅读我们的德国金融指南。+” +# +$ +% +& +’ +( +) +* ++ +, +- +.+/ +0 +1 +2 +3 +4 +5 +6 +7 +8 +9 +: +; +< += +> +?[email protected] +[ +\\u0009+] +^ +_ +` +a +b +c +d +e +f +g +h +i +j +k +l +m +n +o +p +q +r +s +t +u +v +w +x +y +z +{ +| +} +§ +¡ +© +ª +« +Ø +® +° + ± +² +³ +´ +µ +¶ +· +¹ +º +» +¼ +½ +¤ +¿ +× +ß +æ +ð +÷ +ø +þ +đ + ħ +ı +ł + ŋ +œ +ς + ɐ + ɑ + ɒ + ɔ + ɕ + ə + ɛ + ɡ + ɣ + ɨ + ɪ + ɫ + ɬ + ɯ + ɲ + ɴ + ɹ + ɾ + ʀ + ʁ + ʂ + ʃ + ʉ + ʊ + ʋ + ʌ + ʎ + ʐ + ʑ + ʒ + ʔ + ʰ + ʲ + ʳ + ʷ + ʸ + ʷ + ´ + ʾ + ʿ + ˈ + ː + ˡ + ˢ + ˣ + ˤ + α + β + γ + δ + ε + ζ + η + θ + ι + κ + λ + μ + ν + ψ + ο +π + ρ + ς + σ + τ + υ + φ + χ + ψ + ω + а + б + в + г + д + е + ж + з + и + к + л + м + н + о + п + р + с + т + у + ф + х + ц + ч + ш + щ + ъ + ы + ь + э + ю + я + ђ + є + і + ј + љ + њ + ћ + � + � + � + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + ?? + + ?? +� + 带 + 带带 + 带 + ?? + 带 + 带 + ?? + ?? + ?? ى + ه + ه + ä + м + ă + ك + ق + ف + ـ + ٺ + ð + ٸ + ٷ + ٶ + ٵ + ٴ + + ٲ + р + ٰ + ð + ٮ + ك + ق + ف + ـ + ٺ + ư + ٸ + ٷ + ٶ + ٵ + ٴ + + ٲ + р + ٰ + ð + ٮ + ف + ٬ + ٫ + ـ + É + ق + ق + ١ + ٌ + ת + ש + ר + ק + צ + ץ + פ + ף + ע + ס + נ + ן + מ + ם + ל + כ + ך + й + ט + ח + ז + ו + ה + ד + ג + ב + א + ־ +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +� +�

使用 CRISPR 碱基编辑器在 DNA 磁带上进行数字数据存储
摘要:尽管档案数字存储行业已接近其物理极限,但需求却在大幅增长,因此出现了替代产品。最近的努力已经证明了 DNA 作为数字存储介质的巨大潜力,具有卓越的信息耐久性、容量和能耗。然而,大多数提出的系统都需要按需从头 DNA 合成技术,这些技术会产生大量有毒废物,因此不具备工业可扩展性和环保性。受半导体存储设备架构和基因编辑最新发展的启发,我们创建了一种称为“DNA 突变覆盖存储”(DMOS)的分子数字数据存储系统,该系统通过利用组合、可寻址、正交和独立的体外 CRISPR 碱基编辑反应来存储信息,将数据写入绿色合成 DNA 磁带的空白池中。作为概念验证,我们在 DNA 磁带上写下了我们学校徽标的位图表示和本研究的标题,并准确地恢复了存储的数据。

具有不受束缚的逆转录酶和环状 RNA 模板的分裂引物编辑器
Es 可实现删除、插入和碱基替换而不会造成双链断裂 1 。然而,目前的 PE2、PE2* 和 PEmax 效应物(nCas9 与 Moloney 鼠白血病病毒 RT(M-MLV RT)的融合)1 – 3 > 6.3 千碱基 (kb),超出了 AAV 的包装能力。高产量生产如此大的蛋白质或 mRNA(用于核糖核蛋白 (RNP) 或 RNA 递送)也是一项挑战。尽管一些拆分策略已用于递送 Cas9 相关基因组编辑工具 4 ,包括拆分内含肽 5 – 7 和 MS2(参考文献 8 – 10)或 SunTag 11 系链,但大多数拆分方法才刚刚开始应用于 PE 2、12、13。这些元素增加了 PE 系统的尺寸、分子复杂性以及生产和递送负担,并且限制了 PE 开发的组合吞吐量(即核酸酶和 RT 成分的混合和匹配)。pegRNA 优化对于有效的引物编辑也很重要。当前的 pegRNA 是一种结合 RNA,由 sgRNA 和包含 RT 模板 (RTT) 和引物结合位点 (PBS) 的 3′ 延伸组成。尽管在 PE 系统中整合 RNA 分子很简单,但由于 PBS 和间隔区之间不可避免的碱基配对以及潜在的 RTT-支架相互作用,它容易发生 RNA 错误折叠。最后,pegRNA 中的 3′ 末端延伸暴露在外,易受核酸酶降解,这可能会损害 pegRNA 的完整性。虽然 3′ 末端二级结构提高了 pegRNA 的稳定性 14 ,但仍需要进一步努力减少 pegRNA 的错误折叠和不稳定性。

改进的胞嘧啶碱基编辑器的基因组和转录组范围的脱靶分析
1 博伊斯汤普森研究所,纽约州伊萨卡 14853,美国;2 马里兰大学植物科学与景观建筑系,马里兰州帕克分校,美国;3 扬州大学农学院,江苏省作物基因组学与分子育种重点实验室/植物功能基因组学教育部重点实验室,扬州 225009,中国;4 扬州大学江苏省粮食作物现代生产技术协同创新中心,扬州 225009,中国;5 康奈尔大学植物育种与遗传系,纽约州伊萨卡 14853;6 马里兰大学生物科学与生物技术研究所,马里兰州罗克维尔 20850。Ɨ 上述作者对本文贡献相同。