XiaoMi-AI文件搜索系统
World File Search System荧光的

细菌群体传感信号是在全球丰富的emiliania huxleyi
海洋浮游植物,病毒和细菌之间的相互作用驱动生物地球化学循环,塑造海洋营养结构并影响全球气候。微生物产生的化合物已成为影响真核生理学的关键参与者,进而重塑微生物群落结构。这项工作旨在揭示细菌群体传感分子2- heptyl-4-喹诺酮(HHQ),该分子是由海洋γ-帕特罗氏菌杆菌spps spp。生成的,可阻止病毒诱导的cocimopol cocicocipol hh hhh cocicocipocipocipocipocipol hh spp。先前的工作已经建立了烷基素醇作为二羟基脱氢酶(Dhodh)的抑制剂,这是一种基本酶,促进了嘧啶生物合成和潜在的抗病毒药物靶标的第四步。huxleyi dhodh的N末端截断版在大肠杆菌中异源表达,纯化和动力学表征。在这里,我们显示HHQ是E. huxleyi Dhodh的有效抑制剂(2.3 nm的K I)。E。huxleyi细胞暴露于brequinar(典型的人类dhodh抑制剂)中,经历了直接但可逆的细胞停滞,这种作用反映了先前观察到的HHQ诱导的细胞静止。然而,brequinar治疗缺乏在HHQ暴露的赫uxleyi中观察到的其他显着影响,包括细胞大小,叶绿素荧光的显着变化以及免受病毒诱导的裂解的保护,表明HHQ具有尚未发现的尚未发现的生理靶标。一起,这些结果表明细菌群体传感分子在海洋生态系统中三方相互作用中的新颖而复杂的作用,为探索微生物化学信号传导在Algal Bloom调节和宿主肺病动力学中的作用开辟了新的途径。

引用纸浆版本(APA):Cleary,S。R.,Seflova,J.,Cho,E.E.,E.E.,Bisht,K.,Khandelia,H.,Espinoza-Fonseca,L.M。,L. M.,&Robia,S.L。(2024)。
磷兰班(PLB)是一种跨膜小肽,可调节心脏肌肉中的肌质网Ca 2+ -ATPase(SERCA),但这种调节的物理机制仍然很熟悉。PLB降低了活性SERCA的Ca 2+敏感性,从而增加了泵循环所需的Ca 2+浓度。然而,当不存在ATP时,PLB不会降低Ca 2+与SERCA的结合,这表明PLB不会抑制SERCA Ca 2+ afintient。对这些看似冲突的结果的主要解释是,PLB在与Ca 2+结合相关的SERCA酶促循环中的转变减慢了转运Ca 2+的依赖性,而不会实际影响Ca 2+协调位点的等电数。在这里,我们考虑了另一个假设,即在没有ATP的情况下,Ca 2+结合的测量可忽略核苷酸结合的重要变构效应,从而增加了SERCA Ca 2+结合效果。我们推测PLB通过逆转这种同义来抑制SERCA。为了测试这一点,我们使用了荧光的SERCA生物传感器来量化非循环SERCA的Ca 2+在存在和不存在不可用的ATP-ANALOG AMPPCP的情况下。核苷酸激活增加了SERCA Ca 2+的原性,并且通过PLB的共表达逆转了这种效果。有趣的是,在没有核苷酸的情况下,PLB对Ca 2+的原性没有影响。这些结果调解了先前的ATPase分析与Ca 2+结合测定的冲突观察结果。此外,SERCA的结构分析揭示了连接ATP和Ca 2+结合位点的新型变构途径。我们提出的这一途径被PLB结合所破坏。因此,PLB通过通过ATP中断泵的变构激活而降低了SERCA的平衡Ca 2+。因此,PLB通过通过ATP中断泵的变构激活而降低了SERCA的平衡Ca 2+。

对分枝杆菌氧化磷酸化抑制剂的简单测定
氧化磷酸化,电子传输链(ETC)和三磷酸腺苷(ATP)合酶的联合活性已成为抗生素治疗感染毒成菌和相关病原体的抗生素的宝贵靶标。在氧化磷酸化中,ET等建立了跨膜电化学质子梯度,从而为ATP合成提供动力。通过基于荧光素酶的ATP合成或测量氧气消耗的检测来监测氧化磷酸化可能在技术上具有挑战性且昂贵。这些局限性降低了这些方法在表征分枝杆菌氧化磷酸化抑制剂的效用。在这里我们表明,基于荧光的倒膜囊泡酸化(IMV)可以检测和区分抑制ETC的抑制,抑制ATP合酶和非特异性膜解偶联。在该测定中,来自smegmatis的IMV通过ETC或ATP合酶的活性酸化,后者对遗传进行了修饰,以使其充当ATP驱动的质子泵。通过9-氨基-6-氯-2-甲氧基因氨酸的荧光监测酸化,该酸氧化含量会在酸化的IMV中积聚和淬灭。非特异性膜解耦合器可防止琥珀酸酯和ATP驱动的IMV酸化。相比之下,ETC复合物III 2 IV 2抑制剂TelaceBEC(Q203)可防止琥珀酸驱动的酸化,但不能防止ATP驱动的酸化和ATP合酶抑制剂bedaquiline防止ATP驱动的酸化,但不能防止ATP驱动的酸化,但不能防止琥珀酸助长驱动的酸化。我们使用该测定法表明,正如先前提出的那样,兰索拉唑硫化物是复合物III 2 IV 2的抑制剂,而硫代嗪则是非特定于分枝杆菌膜的抑制剂。总体而言,该测定是简单,低成本且可扩展的,这将使其可用于识别和表征新的分枝杆菌氧化磷酸化抑制剂。

SURF 和 VURP 摘要将最小浸入与...联系起来
将曲面上扁平线束的最小浸入与临界特征值度量联系起来 Santiago Adams 导师:Antoine Song 在现有文献中,第一个特征值在曲面上临界的度量与该曲面在任意维球面中的最小浸入之间存在着密切的联系。我们知道,对于具有临界度量的曲面,存在一组拉普拉斯算子的特征函数,它们定义了进入球面的最小浸入。我们旨在使用局部参数将该理论扩展到扁平线束特征截面的情况。也就是说,给定一个第一个特征值在线束上临界的度量,我们旨在使用其特征截面的升力来定义其通用覆盖在球面中的最小浸入,并更好地理解是否存在原始曲面进入球面的最小浸入。伊辛铁磁体在经典和量子极限下的热力学性质 Sophia Adams 导师:Thomas Rosenbaum 和 Daniel Silevitch 该项目旨在探测模型伊辛铁磁体 LiHoF 4 在经典和量子相变中的热力学性质。经典跃迁发生在临界温度 1.53 K 和零磁场下,而量子跃迁发生在零温度极限下 50 kOe 量级的临界横向磁场下。我们将使用比热数据来比较两个跃迁的临界指数及其之间的交叉。 一种使用基于分类器的生成器生成和预筛选蛋白质以确定结合亲和力的新方法 Victoria Adams 导师:Matt Thomson 和 Alec Lourenco 由于当前方法筛选蛋白质结合功效的速度和规模,测试新的工程结合蛋白设计非常无效。定量而不是定性筛选新蛋白质将进一步提高效率。 Thomson 实验室开发了一种高通量筛选方法,用于收集有关结合蛋白的信息并实现蛋白质设计。在我的项目中,我致力于开发一种使用蛋白质语言模型预筛选生成蛋白质的新方法。应用现有的蛋白质大型语言模型 (pLLM),例如进化尺度模型 (ESM) 和 AlphaFold 2 & 3,我正在研究一种生成蛋白质然后预筛选其结合亲和力的方法。我还有机会学习如何使用实验室的高通量筛选分析来实验性地测试蛋白质设计。到目前为止,我还没有完全开发的方法/模型,但我有一个需要微调的基本分类器,并且需要一个仍需要指定最佳参数的生成器。我希望能够完成这些编程改进,并可能能够在夏季结束前通过应用高通量筛选来测试它们。来自路径积分的时间类纠缠 Zofia Adamska 导师:John Preskill 和 Alexey Milekhin 大多数量子力学形式主义都从不同的角度来看待空间和时间,这从相对论物理学的角度来看似乎是不自然的。为了解决这种不对称性,我们提出了一种时空密度矩阵的新定义,该定义源自路径积分方法,以更好地分析时空中的量子信息。我们的动机基于相对论量子场论中的观察,其中该密度矩阵的 Renyi 熵与通过从空间类分离到时间类分离的解析延续得出的结果完全一致。我们演示了如何使用这个密度矩阵来限制时空相关函数,并表明我们的界限比其他方法更紧并且遵循 Lieb-Robinson 界限。此外,我们在量子计算机上测试了这个时空密度矩阵对单量子比特系统的预测。使用我们的方法计算的时空纠缠构成了热化的新探针,并且可以为选择用于量子多体系统时间演化的有效张量网络假设提供启示。使用合成细胞建立病毒宿主相互作用的最小模型 Layla Adeli 导师:Richard Murray 和 Zach Martinez 利用最小模型研究合成细胞病毒感染的潜力使其成为研究尚未得到充分研究的病原体的首选。为了设计 PhiX174 噬菌体的合成宿主,我们尝试将 PhiX174 识别的脂多糖 (LPS) 整合到脂质体膜中,以潜在地封装无细胞转录、翻译和复制系统 (PURE Rep)。此外,设计为在脂质体内由 PhiX174 基因触发时发出荧光的立足点开关可以检测 PhiX174 基因组的 DNA 转录——我们目前的工作包括设计一种具有高效性的开关。我们已经成功生产出脂质体,并正在努力整合检测机制我们在量子计算机上测试该时空密度矩阵对单量子比特系统的预测。使用我们的方法计算的时空纠缠构成了一种新的热化探测,可以为选择一种有效的张量网络假设来研究量子多体系统的时间演化。使用合成细胞建立病毒宿主相互作用的最小模型 Layla Adeli 导师:Richard Murray 和 Zach Martinez 利用最小模型研究合成细胞病毒感染的潜力使合成细胞成为研究尚未得到充分研究的病原体的首选。为了设计 PhiX174 噬菌体的合成宿主,我们尝试将 PhiX174 识别的脂多糖 (LPS) 整合到脂质体膜中,以潜在地封装无细胞的转录、翻译和复制系统 (PURE Rep)。此外,当脂质体中的 PhiX174 基因触发时,设计为发出荧光的立足点开关可以检测 PhiX174 基因组的 DNA 转录——我们的工作目前包括设计一种具有高效性的立足点开关。我们已经成功生产出脂质体,并正在努力整合检测机制我们在量子计算机上测试该时空密度矩阵对单量子比特系统的预测。使用我们的方法计算的时空纠缠构成了一种新的热化探测,可以为选择一种有效的张量网络假设来研究量子多体系统的时间演化。使用合成细胞建立病毒宿主相互作用的最小模型 Layla Adeli 导师:Richard Murray 和 Zach Martinez 利用最小模型研究合成细胞病毒感染的潜力使合成细胞成为研究尚未得到充分研究的病原体的首选。为了设计 PhiX174 噬菌体的合成宿主,我们尝试将 PhiX174 识别的脂多糖 (LPS) 整合到脂质体膜中,以潜在地封装无细胞的转录、翻译和复制系统 (PURE Rep)。此外,当脂质体中的 PhiX174 基因触发时,设计为发出荧光的立足点开关可以检测 PhiX174 基因组的 DNA 转录——我们的工作目前包括设计一种具有高效性的立足点开关。我们已经成功生产出脂质体,并正在努力整合检测机制

通过CRISPR-CAS靶向裂开志贺毒素基因的靶向裂解在动物模型中
摘要。收音机和手机使用振荡载体信号的频率调制(FM)来可靠地传输多路复用数据,同时拒绝噪声。在这里,我们使用遗传编码的蛋白振荡器(GEOS)作为电路中的载波信号来建立该范式的生化类似物,以实现单细胞数据的连续实时FM流。GEOS是由进化多样的思想家庭ATPase和激活因子模块构建的,这些模块在人类细胞中共表达时会产生快速的合成蛋白振荡。这些振荡用作单细胞载体信号,频率和振幅由GEO组件水平和活动控制。我们系统地表征了169个ATPase/Activator Geo对和具有多个竞争激活剂的工程师复合GEO,以开发一个用于波形编程的全面平台。使用这些原理,我们设计了对细胞活性调节地理频率的电路,并使用校准的机器学习模型解码其响应,以证明单个单元中转录和蛋白酶体降解动力学的敏感,实时FM流。GEOS建立一个动态控制的生化载体信号,解锁抗噪声的FM数据编码范式,为动态单细胞分析开辟了新的途径。简介。细胞动态调节不同时间尺度的基因表达,蛋白质定位和信号传导状态,以执行必不可少的生物学功能1-4。虽然基因组,转录组和蛋白质组学方法可以提供单细胞态5-8的快照,但实时遵循单个细胞的轨迹的能力对于理解动态细胞和生物体行为如何编码和功能1,9,10至关重要。这些单细胞动力学通常是使用荧光记者在显微镜下进行跟踪的,其强度或定位为您感兴趣的数据提供了代理10-16。虽然功能强大,但这些工具对扩展单细胞动力学和数据聚合的扩展跟踪构成了挑战,因为任意信号强度在仪器上各不相同,并且对光漂白和噪声17敏感。此外,传统基于荧光的工具生成的信号缺少元数据来识别信号的基本细胞来源,从而使密集的细胞环境中重叠信号的分离变得困难。
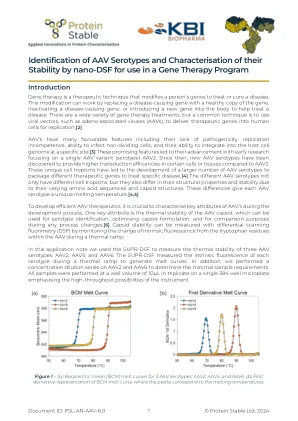
识别AAV-Serotypes and-thacteration-of-...
基因治疗是一种治疗技术,可修饰一个人的基因治疗或治愈疾病。这种修饰可以通过用健康的基因副本代替引起疾病的基因,使疾病的基因失活或将新基因引入体内以帮助治疗疾病。有多种基因疗法治疗,但是一种常见的技术是使用病毒载体,例如腺相关病毒(AAV),将治疗基因传递到人类细胞中进行复制。[2] AAV具有许多有利的特征,包括缺乏致病性,复制无能力,感染非分散细胞的能力以及在特定部位中整合到宿主细胞基因组中的能力。[3]这些有前途的特征导致了它们的进步,早期研究重点是单个AAV变体(血清型)AAV2。从那时起,与AAV2相比,已经发现了新的AAV血清型在某些细胞或组织中提供较高的转导效率。这些独特的细胞偏向主义已导致大量AAV血清型的发展来包装不同的治疗基因来治疗特定疾病。[4]不同的AAV血清型不仅具有不同的细胞向量,而且由于其不同的氨基酸序列和衣壳结构,它们的结构特性和稳定性也有所不同。这些差异使每种AAV血清型都具有独特的熔化温度。[4,5]为了开发有效的AAV疗法,在开发过程中表征AAV的关键属性至关重要。SUPR-DSF在热坡道期间测量了每种血清型的固有荧光,以产生熔体曲线。一个关键属性是AAV CAPSID的热稳定性,可用于血清型识别,优化衣壳配方以及在任何过程更改中进行比较。[6]可以通过在热坡道期间从AAV中的色氨酸残基中监测固有荧光的变化,可以通过差异扫描荧光法(DSF)测量衣壳稳定性。在此应用程序注释中,我们使用SUPR-DSF来测量三种AAV血清型的热稳定性:AAV2,AAV5和AAV6。此外,我们在AAV2和AAV6上进行了浓度稀释系列,以确定最小样品要求。所有样品均以10µl的井体积为10µl,在单个384孔微孔板上一式三份,强调该仪器的高通量可能性。

聚腺苷二磷酸的动力学和静态分析
聚腺苷二磷酸核糖聚合酶 (PARP) 蛋白家族参与多种功能,最显著的是 DNA 损伤反应。癌症易受 DNA 损伤的影响,这导致了几种 PARP 抑制剂 (PARPi) 的开发。这类药物已被证明对卵巢癌、乳腺癌和前列腺癌有治疗效果,但反应各不相同。因此,诊所需要选择可能从这些靶向疗法中受益的患者。体内 18 F-氟邻苯二甲酸酯摄取成像已被证明与组织中的 PARP-1 表达相对应。本研究表征了 18 F-氟邻苯二甲酸酯的药代动力学,并测试了动力学和静态模型,以指导未来研究中评估 18 F-氟邻苯二甲酸酯作为 PARPi 治疗反应生物标志物的指标选择。方法:14 名前瞻性入选的卵巢癌患者注射 18 F-氟苯那敏,注射后进行 60 分钟动态成像,随后进行最多 2 次全身扫描,测量静脉血活性和代谢物。从动态图像和全身扫描中提取 SUV 最大值和 SUV 峰值。评估动力学参数估计值和 SUV 与组织 PARP-1 免疫荧光的相关性(n=7)。群体动力学参数的模拟可以估计测量偏差和参数估计的精度。结果:18 F-氟苯那敏血液清除率各不相同,但不同患者的标记代谢物谱相似,支持使用群体母体分数曲线。可逆性2组织室模型和Logan参考组织分布体积(DVR)在PET采集的第一个小时内的总分布体积与免疫荧光检测的肿瘤PARP-1表达相关(分别为r=0.76和0.83;P=0.05)。DVR偏差和精度估计分别为6.4%和29.1%。从中点为57.5、110±3和199±4分钟的图像获得的SUV max和SUV peak与PARP-1表达高度相关(平均值±SD,r=0.79;P=0.05)。结论:注射后55-60分钟及以后的肿瘤SUV max和SUV peak以及至少60分钟的DVR似乎是PARP-1结合的可靠非侵入性测量方法。 18 F-氟苯那敏在卵巢癌中的吸收最好用可逆结合模型来描述。然而,示踪剂吸收的药代动力学模式有些变化,尤其是在后期。

质子泵抑制剂(PPIS)的Jurkat T淋巴细胞凋亡诱导
凋亡(通常称为程序性细胞死亡)不断发生在人类中。随着癌细胞的酸度增加,诱发了凋亡。在健康细胞中,质子泵蛋白允许H +离子渗透到细胞膜,从而调节pH值。然而,质子泵抑制剂(PPI),例如奥美拉唑,防止质子运动,导致pH调节。在先前的研究中,奥美拉唑诱导了Jurkat T淋巴细胞的细胞死亡;但是,尚无证实细胞是通过细胞凋亡或通过坏死而死亡的,而细胞爆发。通过使用膜联蛋白-V染色,可以测量奥美拉唑,右氯唑唑和埃索美吡唑对凋亡诱导的影响。细胞死亡。右兰索拉唑和埃索美拉唑在18小时时均达到100%的凋亡,表明它们具有较早的凋亡激活点。为了测量细胞活力的程度,通过用小钙蛋白 - 乙酰氧基甲基(AM)染料染色细胞来测量胞质酯酶活性。Jurkat细胞暴露于Omeprazole,Dexlansoprazole和Esomeprazole六个小时,并监测30小时以测量生存能力。阿霉素是一种已知的化学治疗性,在测试凋亡诱导和生存力时也被用作阳性对照。使用荧光显微镜成像时,由于膜联蛋白V-FITC的结合而导致凋亡荧光的任何细胞以及由于PI的结合而导致的坏死细胞荧光。用钙软蛋白AM(如果细胞荧光,它们)被认为是可行的,而非荧光细胞被认为是坏死的。在30小时的标记下,右倾角唑的生存力最小(40.0±3.5%的细胞可行),其次是阿霉素(62.9±1.8%),埃索美普唑(66.2±1.6%)和欧洲普拉唑(69.29±2.01%)(69.29±2.01%),在比较(71%)中(71%)(71%)。右兰索拉唑的生存能力低,表明需要使用相同的PPI和暴露方法进行毒性研究,以确定最佳药物浓度。奥美拉唑和埃索美瑞唑的最佳浓度为1 µm,右兰索拉唑啉为0.5 µm。未来的研究包括使用膜联蛋白V-FITC和碘化丙啶(PI)染料在确定浓度下测试细胞死亡方法。

编制螺旋钻电子放射性药物治疗的潜力
对编辑者:我们非常感兴趣地阅读了《命运 - 布莱斯特04试验的结果》,“莫迪等人的trastuzumab deruxtecan trastuzumab deruxtecan”,莫迪等人。并发表在《新英格兰医学杂志》(1)上。在美国临床肿瘤学会议上,这些令人印象不佳2022。我们坚信核医学界应该熟悉这项关键临床试验的结果,该试验正在为HER2靶向的宠物成像开辟新的令人兴奋的前景。HER2是由ERBB2癌基因编码的膜蛋白。ERBB2在10% - 15%的浸润性乳腺癌中得到扩增,并分类为HER2阳性用于临床实践。通过免疫组织化学鉴定这些肿瘤均被评分为3 1或2 1,并且具有通过原位杂交评估的基因扩增。到目前为止,只有Her2阳性乳腺癌可以由抗HER2疗法靶向。 其余的肿瘤没有可检测到的HER2或低水平,分别称为“ Her2-Zero”和“ Her2-low”患者。 在Destiny-Breast04中,T-DXD与HER2-LOW MBC患者的标准化学疗法相比,T-DXD提供了具有临床意义的好处。 我们正在重新思考评估HER2身份的方式。到目前为止,只有Her2阳性乳腺癌可以由抗HER2疗法靶向。其余的肿瘤没有可检测到的HER2或低水平,分别称为“ Her2-Zero”和“ Her2-low”患者。在Destiny-Breast04中,T-DXD与HER2-LOW MBC患者的标准化学疗法相比,T-DXD提供了具有临床意义的好处。我们正在重新思考评估HER2身份的方式。t-dxd成功延长了Her2-low MBC患者(1)患者的无进展生存期和整体存活率。与化学疗法组中中位无进展的生存期相比,T-DXD组的患者的中位前进展生存期为10 mo,因此疾病进展或死亡的风险降低了50%(危险比,0.50; 95%CI,0.40 - 0.63; p,0.0001)。发现必定会改变高级乳腺癌分类的方式。然而,当前的标准免疫组织化学测定未标准化以区分Her2-low与Her2-Zero患者(2),而是识别HER2阳性患者。病理学家目前正在修改其解释染色的方法,以准确识别HER2-LOW患者。问题仍然开放,目前正在研究的基于免疫荧光的新策略还是正在研究的质谱法的制定是否会改善患者的HER2状态评估(3)。在转移性环境中,转移部位之间的HER2状态异质性在评估中仍然具有挑战性,并且已知会影响对疗法的反应(4)和临时肿瘤内HER2异质性。her2宠物似乎是一种有希望的替代工具,用于定量和无创地监测全身HER2表达。尽管大多数发表的研究主要集中在HER2阳性患者(5,6),但Ulaner等人的关键论文。出版于

实现特定 RNA 靶向和降解的新型化学方法:应用于 SARS-CoV-2 RNA
当前药物化学面临的主要挑战之一是使用小分子药物靶向非编码 RNA。与传统的药物发现方法不同,它需要改变视角才能取得成功。正如 Disney 及其同事 1 在本期《ACS Central Science》中所描述的那样,针对 SARS-CoV-2 RNA 的特异性配体的发现、核糖核酸酶靶向嵌合体 (RIBOTAC) 的开发以及使用尖端化学生物学工具对细胞内作用机制的详细研究,凸显了新化学模式可能对未来疗法产生的影响。这项工作的应用领域涉及针对导致当前全球健康危机的病毒 SARS-CoV-2。人们在药物重新定位、大规模筛选和疫苗开发方面做出了许多努力;然而,显然需要研究用于创新治疗方法的新靶点和新生物活性分子,以增加抗病毒药物库。RNA 被认为是基因表达过程中的中间体,现在是药物发现的有效靶点。 2 RNA 是一种大分子,参与转录、翻译和基因表达调控等重要生物过程。事实上,非编码 RNA 表达和/或功能的许多失调与许多病理直接相关,例如神经系统疾病、心脏病或癌症。最值得注意的是,超过 70% 的人类基因组在非编码 RNA 中转录,而只有 1.5% 编码蛋白质。由于这些蛋白质中只有一小部分代表上市药物的实际靶标,因此很明显,将非编码 RNA 添加到潜在治疗靶标将大大增加药物开发的前景。由新型 RNA 病毒 SARS-CoV-2 引起的冠状病毒大流行凸显了 RNA 靶向疗法在治疗 RNA 病毒引起的感染方面的潜力。这些疗法之前曾在文献中用于 HIV、HCV 或流感病毒。自 20 世纪 40 年代第一批 RNA 配体作为抗生素进入市场以来,新的 RNA 靶向药物的发现取得了重大进展,例如氨基糖苷类或四环素类,以及最近的恶唑烷酮类。3 近年来,RNA 结合剂的进展包括不同类型的原创、有时不寻常的药物发现策略。还开发了用于识别非常特异性 RNA 配体的先导识别策略,例如 Inforna。4 后者是一种基于二维组合筛选 (2-DCS) 和通过测序 (StARTS) 实现的结构-活性关系相结合的技术。将这种统计方法应用于筛选结果可以预测 RNA 库成员的亲和力和选择性并评分结合相互作用。4 该方法已被证明非常有效,可以成功预测特定 RNA 二级结构的化合物。各种筛选技术(例如微阵列或基于荧光的检测)也已用于发现 RNA 配体以及基于结构的选择性配体设计。5-7