XiaoMi-AI文件搜索系统
World File Search System轴突

大脑皮层锥体神经元的计算能力
皮质锥体神经元的电活动由结构稳定、形态复杂的轴突树突树支持。轴突和树突在长度或口径方面的解剖差异反映了神经信息输入或输出的底层功能特化。为了正确评估锥体神经元的计算能力,我们分析了 NeuroMorpho.Org 数据库中的大量三维数字重建数据集,并量化了小鼠、大鼠或人类大脑皮层不同区域和层次的基本树突或轴突形态测量值。根据获得的形态测量数据对参与神经元电脉冲的离子总数和类型的物理估计,结合活动大脑消耗的葡萄糖所驱动的神经递质释放和信号传导的能量,支持在热力学允许的 Landauer 极限下进行高效的大脑计算,从而实现不可逆的逻辑运算。电压感应 S4 蛋白 Na + 、K + 或 Ca 2+ 离子通道 α 螺旋中的单个质子隧穿事件非常适合用作单个 Landauer 基本逻辑运算,然后通过穿过开放通道孔的选择性离子电流进行放大。这种计算门控的小型化允许在人类大脑皮层中每秒执行超过 1.2 个 zetta 逻辑运算,而不会因释放的热量而燃烧大脑。

用于拓扑的自监督边缘检测重建...
许多基于机器学习的轴突追踪方法依赖于带有分割标签的图像数据集。这需要领域专家的手动注释,这需要大量劳动力,并且不适用于以细胞或亚细胞分辨率对半球或整个脑组织进行大规模脑映射。此外,保留轴突结构拓扑对于理解神经连接和大脑功能至关重要。自监督学习 (SSL) 是一种机器学习框架,允许模型在未注释的数据上学习辅助任务,以帮助完成监督目标任务。在这项工作中,我们提出了一种新颖的 SSL 辅助任务,即为面向拓扑的轴突分割和中心线检测的目标任务重建边缘检测器。我们使用小鼠大脑数据集对三个不同的 SSL 任务进行了 3D U-Nets 预训练:我们提出的任务、预测排列切片的顺序和玩魔方。然后,我们在不同的小鼠大脑数据集上评估了这些 U-Nets 和基线模型。在所有实验中,针对我们提出的任务进行预训练的 U-Net 分别将基线的分割、拓扑保留和中心线检测提高了 5.03%、4.65% 和 5.41%。相比之下,切片排列和魔方预训练的 U-Net 并没有比基线有持续的改进。

神经系统的组织和功能第 3 章
• 前脑 • 中脑 • 后脑 • 3 周:大小均等 • 11 周:前脑成为最大部分 • 下管成为脊髓 • 灰质由无髓鞘细胞体组成。白质是神经元,其轴突有髓鞘。

审查细胞信号传导,生长因子...
摘要____________________________________________________________________________________________________________________________构成了整个神经系统。由创伤,事故和其他相关因素引起的周围神经损伤总是会导致感觉和运动功能均大大丧失。可以通过重建功能轴突成功恢复受伤的神经。尚未优化PNI的完整恢复。外源生长因子(GF)是一种可用于神经再生的新治疗策略。生长因子作用机理基于通过与单个受体结合激活信号级联的能力,以发挥多种作用并恢复神经元和组织再生。尽管GFS受到短暂的半衰期和快速失活的限制。神经导管的使用能够减少这些限制。神经管道是良好的生物相容性和生物功能。关键字:轴突,生长因子,周围神经损伤,信号级联。

在健康和疾病中的体内映射脑微观结构...
扩散磁共振成像在脑白质中对组织微观结构的体内敏感性独特,在发育过程中经历了重大变化,并且在几乎every的神经系统疾病中受到损害。然而,面临的挑战是开发针对人类MRI扫描中几分钟的细胞特征的生物标志物。在这里,我们量化了多区域扩散建模框架对轴突的密度,方向和完整性的灵敏度和特异性。我们证明,使用基于机器学习的估计量,我们的生物物理模型捕获了轴突在早期发育,急性缺血和多发性硬化症中的文化变化(总n = 821)。微观结构映射的方法论广泛适用于临床环境和大型成像联盟数据,以研究发展,衰老和病理学。

岩土工程中的人工智能
1 人工智能的概念 人工智能 (AI) 是软件应用程序和服务基于决策算法 (ADM) 模仿认知思维和智能行为的能力。具有人工智能的软件应用程序也称为 ADM 系统。为了开发允许人工智能系统基于认知思维和智能行为做出决策的算法,需要理解智能、思维和学习的本质。1949 年,唐纳德·赫布 (Donald Hebb) 提出了学习规则 (HEBB 1950)。学习规则认为思维与处理信息所需的神经活动有关,描述了这些活动对神经元之间的连接和神经网络上的突触可塑性的影响。为了处理信息,神经元使用来自不同树突的所有输入信号来形成输出信号,该输出信号通过轴突发送到连接的神经元。轴突的频繁使用会加强神经元之间的连接,而不使用则可能导致连接和轴突的缺失。强连接有利于知识的恢复。因此,学习旨在为相关知识建立强连接(见图 1)。赫布规则是人工智能概念发展的关键发现。当“人工智能”一词于 1956 年在达特茅斯研讨会上首次使用时,研究重点是寻找在可实现算法中表示知识的形式化。因此,许多科学领域都参与了人工智能概念的发展:

早期炎症通过上调 IL-1 b 导致体内神经回路形成失调
发育过程中的神经免疫相互作用与神经发育障碍的发病机制密切相关,但导致神经回路失调的机制尚不清楚。我们对斑马鱼幼鱼正在发育的视网膜顶盖系统进行了体内成像,以表征免疫系统激活对原型感觉处理回路细化的影响。急性炎症损伤诱导了鱼幼鱼正在发育的视网膜轴突的超动态重塑,并在几天内增加了轴突树突的精细化。使用 GCaMP6s 转基因鱼中的钙成像,我们发现这些形态变化伴随着顶盖细胞视力下降的转变。视觉引导行为任务中较差的表现支持了这一发现。我们进一步发现,促炎细胞因子白细胞介素-1b (IL-1b) 因炎症刺激而上调,而 IL-1b 的下调则消除了炎症对轴突动力学和生长的影响。此外,IL-1b 变形动物的视网膜神经节细胞树突基线分支与对照幼虫有显著差异,并且它们在捕食试验中的表现受损,表明该细胞因子在正常神经元发育中发挥作用。这项研究建立了一个简单而有效的非哺乳动物发育免疫激活模型,并证明了 IL-1b 在介导炎症对神经元回路发育的病理影响方面的作用。

大脑中是否有光通信通道?
尽管在神经科学方面取得了长足的进步,但仍然存在有关大脑的基本问题,包括主观经验和意识的起源。一些答案可能依赖于新的物理机制。鉴于在大脑中发现了生物光子,探索神经元除了使用精心研究的电信号外使用光子通信很有趣。大脑中的这种光子通信需要波导。在这里,我们回顾了最近的工作(S. Kumar,K。Boone,J。Tuszynski,P。Barclay和C. Simon,Scientific Reports 6,36508(2016)),建议髓鞘轴突可以用作光子波导。考虑到其现实的缺陷,对髓鞘轴突中的光传递进行了建模,并在体内和体外提出了实验,以检验该假设。讨论了对量子生物学的潜在影响。

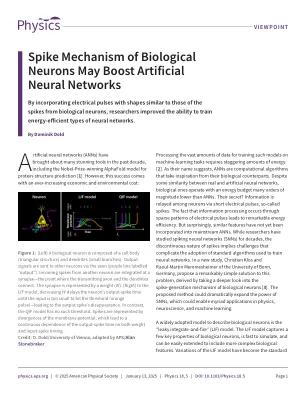
生物神经元的尖峰机制可能会促进人工神经网络
图1 :(左)生物神经元由细胞体(三角形结构)和树突(小分支)组成。输出信号通过轴突(标记为“输出”)发送到其他神经元。来自另一个神经元的传入尖峰集成在突触处,即传输轴突和树突连接的点。突触由重量(W)表示。(右)在LIF模型中,降低W会延迟神经元的输出尖峰时间,直到输入太小而无法达到阈值(橙色脉冲)(橙色脉冲),并导致输出尖峰的消失。相反,QIF模型没有这样的阈值。尖峰由膜电位的差异表示,这导致输出跨度时间对重量和输入跨度的时机的连续依赖性。信用:D。Dold/维也纳大学;由APS/ Alan Stonebraker div>改编