XiaoMi-AI文件搜索系统
World File Search System辅助因子

针对POU2F-POU2AF转录因子驱动的MSWI/SNF复合物
总结POU2F3-POU2AF2/3(OCA-T1/2)转录因子复合物是簇状细胞谱系和簇状细胞样小细胞肺癌(SCLC)的主调节剂。在这里,我们发现SCLC(SCLC-P)的POU2F3分子亚型表现出对哺乳动物开关/不可发酵(MSWI/SNF)染色质重塑复合物的活性的精致依赖性。SCLC-P细胞系对MSWI/SNF ATP酶蛋白水解靶向降解剂的纳摩尔水平敏感。pou2f3及其辅助因子与MSWI/SNF复合物的组件相互作用。POU2F3转录因子复合物在MSWI/SNF ATPase降解时从染色质中驱逐,从而导致SCLC-P细胞中下游致癌信号传导的衰减。一种新型的,可生物利用的MSWI/SNF ATPase Protac Degrader,AU-24118,相对于SCLC-A亚型,SCLC-P中表现出优先效率,并且在旋界模型中显着降低了肿瘤的生长。AU-24118没有改变肺或结肠中正常的簇状细胞数,也没有在小鼠中表现出毒性。b细胞恶性肿瘤对POU2F1/2辅助因子POU2AF1(OCA-B)的依赖性也对MSWI/SNF ATPase ATPase降解非常敏感。从机械上讲,在多个骨髓瘤细胞压实的染色质,移位POU2AF1和IRF4的MSWI/SNF ATP酶降解器处理中,并降低了IRF4信号传导。与POMALIDOMIDE相比,在POU2AF1依赖性的多发性骨髓瘤的鼠模型中,AU-24118增强了生存率,Pomalidomide是多发性骨髓瘤的批准治疗。综上所述,我们的研究表明,POU2F-POU2AF驱动的恶性肿瘤对MSWI/SNF复合物具有内在的依赖,代表了治疗性脆弱性。关键字POU2F3,POU2AF1/2/3,MSWI/SNF复合物,Smarca2/4,蛋白水解靶向嵌合体(Protac),小细胞肺癌(SCLC),多发性骨髓瘤,IRF4,IRF4

建议引用:Fehsel,K。(2023)。为什么铁缺乏症/贫血与阿尔茨海默氏病及其合并症有关,如何预防? [ON
摘要:在许多疾病中越来越多地观察到铁代谢受损,但是仍然缺乏对改变铁代谢的细胞影响的更深入的机械理解。此外,描述了阿尔茨海默氏病(AD)及其合并症(如肥胖,抑郁症,心血管疾病和2型糖尿病)的合并症,描述了由于葡萄糖进口减少而导致的神经元能量代谢的遗嘱。这篇综述的目的是介绍两个观察结果之间的分子联系。不足的细胞葡萄糖摄取诱导者增加了铁蛋白的表达,从而导致细胞不含铁池的耗竭并稳定缺氧诱导的因子(HIF)1α。该转录因子诱导葡萄糖转运蛋白(GLUT)1和3的表达,并将细胞代谢转移到糖酵解。如果这条防线不足以足以满足能力的葡萄糖供应,则进一步减少细胞内铁池会影响线粒体电子传输链的酶,并激活AMP激活的激酶(AMPK)。该酶触发了GLUT4向质膜的转运以及细胞成分的自噬回收,以动员能量资源。此外,AMPK激活了铁蛋白噬菌的自噬过程,该过程提供了急需的铁作为辅助因子,作为辅助因子,以合成血红素和铁 - 硫蛋白的合成。该途径的过度激活以铁铁作用而结束,这是一种特殊的铁依赖性细胞死亡形式,而阻碍AMPK激活稳步减少了铁池,导致脾脏和肝脏中铁隔离性低铁血症。持久的铁耗尽会影响红细胞生成,并导致慢性疾病的贫血,这是AD患者及其合并症的常见状况。应施用改善能源供应和细胞葡萄糖摄取的药物,饮食或植物化学物质,以抵消慢性疾病的低铁和贫血。

对核自旋扩散的理论检查...
微秒相干时间在供体的自旋动力学计算中预测 - 受体电子旋转对PÞA 1A在光系统I(PSI)的光激发后创建。研究了由于各向异性蛋白环境对预测的相干时间T m而引起的核自旋扩散(NSD)的影响。紧密定位的对位于电子旋转的位置5 - 8°A的质子的三元组和三元质子显示为在很大程度上控制T m。对PSI晶体结构的了解允许进行自旋动力学计算,其中去除或替换了特定的辅助因子和氨基酸残基,并且鉴定了控制电子脱碳的各向异性环境特征。最后,我们表明单独的NSD无法解释> 3个较短的实验观察到的相干时间,并暗示关键蛋白质位点的甲基可能解释了这种差异。


增加对免疫检查点抑制剂的反应增加了饮食中的蛋氨酸限制
甲烷古细菌是厌氧消化1中的甲烷生产商,它们是电力到气体过程中的生物催化剂2,它们是全球碳循环3中的重要参与者,甚至由天体生物学家研究4,5。氢化甲烷剂使用还原性乙酰-COA途径进行CO 2固定6,这是一种从CO 2合成有机碳的能节能途径和与乙蛋白酶中存在的途径相似的氢。然而,乙酸7与ATP与离子梯度投资的CO 2降低的甲烷生成方式之间存在细微的差异,并且在辅助因子利用率8方面存在差异8。因此,建议同时考虑乙蛋白原和甲烷剂,作为可能的宿主生物,用于从CO 2作为碳源生产燃料和化学物质。

DNA依赖性腺病毒基因A ...
摘要我们已经开发了一种无细胞的系统,用于研究哺乳动物细胞中mRNA的合成。该系统由透析和浓缩的全细胞提取物组成,从HeLa细胞,小分子和转录所需的辅助因子和外源添加DNA组成。RNA聚合酶II的准确介绍完全取决于添加含有启动子的真核DNA。在最佳DNA和提取浓度下,易于检测到来自腺病毒血清型2后期启动子的转录起始,并且可以使用超过4000个核苷酸的特定转录本。在体外合成的RNA包含与体内transkipt相同的5'限制RNase T1 Undeclepleotide。RNA合成还可以在早期和中间腺病毒启动子位点准确地启动。

E2F2 在癌症进展中的作用及其作为治疗靶点的价值
E2F 转录因子家族在调节细胞周期进程和细胞增殖方面起着至关重要的作用。越来越多的证据表明,E2F2 的异常表达或激活是恶性肿瘤中的常见现象。E2F2 已成为各种类型肿瘤发展和进展的关键因素。大量研究证实,E2F2 可能有助于增强肿瘤细胞增殖、血管生成和侵袭性。此外,E2F2 通过与一系列辅助因子和下游靶标(包括细胞凋亡和 DNA 修复)相互作用,对无数细胞过程产生影响。致癌作用中 E2F2 的失调可能归因于多种机制,包括上游调控元件的修改或表观遗传改变。本综述探讨了E2F2在癌症进展中的作用以及针对该致癌途径的既定和新兴的治疗策略,同时也为进一步研究E2F2的生物学功能和临床应用提供了坚实的基础。

电气科学学院(电子与通信工程)基础科学学院(数学)Q-Exam主题&...
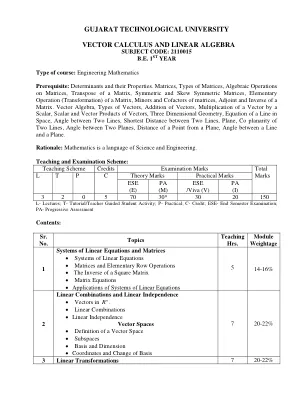
教学大纲:矢量空间,场,子空间,碱基和维度;线性方程,矩阵,等级,高斯消除系统;线性变换,矩阵,rank-nullity定理,二元性和转置的线性变换表示;决定因素,拉普拉斯膨胀,辅助因子,伴随,cramer的规则;特征值和特征向量,特征多项式,最小多项式,Cayley-Hamilton定理,三角剖分,对角线化,有理规范形式,约旦规范形式;内部产物空间,革兰氏阴性正统计,正交投影,线性功能和伴随,遗传学,自我伴随,单一和正常运算符,正常运算符的光谱定理;瑞利商,最小最大原则。双线性形式,对称和偏斜的双线性形式,实际二次形式,西尔维斯特的惯性定律,正定性。

癌症中的营养表观遗传学
营养表观遗传学是指饮食在不改变 DNA 序列的情况下对基因表达的影响。饮食通过影响表观遗传因子的活性和向靶位点的募集来调节表观遗传事件,例如 DNA、RNA 和组蛋白修饰(图 1)。饮食可以促进代谢过程,产生表观遗传因子发挥作用所需的辅助因子,或提供直接结合和调节这些因子活性的分子。此外,饮食会影响转录因子的活性,从而影响表观遗传因子向基因组的募集。重要的是,用小合成分子靶向几种表观遗传因子是当前治疗某些癌症的策略。因此,饮食在癌症发生和发展中的一些有益作用可能是通过调节表观遗传机制来介导的。在本文中,我们将回顾目前用于治疗癌症的治疗靶点的有前景的表观遗传因子,以及饮食产品在调节其活性方面的潜在影响。