XiaoMi-AI文件搜索系统
World File Search System锌指
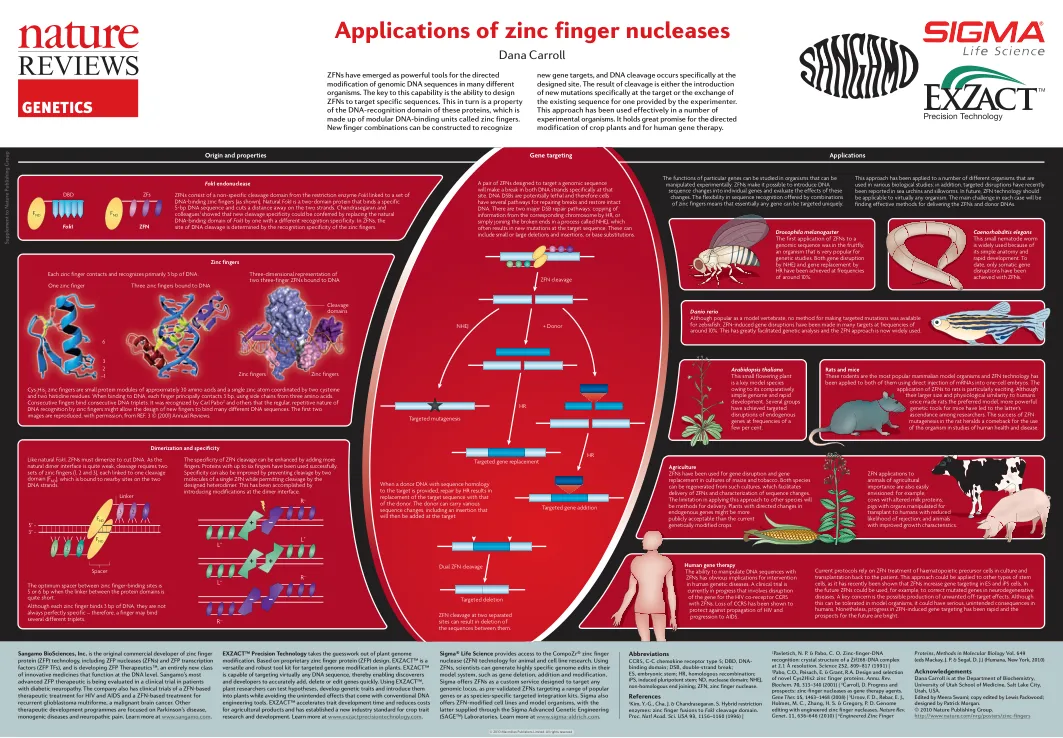
锌指核酸酶的应用
EXZACT™ 精准技术消除了植物基因组改造中的猜测。EXZACT™ 基于专有的锌指蛋白 (ZFP) 设计,是一种多功能且强大的工具包,可用于对植物进行靶向基因组改造。EXZACT™ 能够针对几乎任何 DNA 序列,从而使发现者和开发者能够快速准确地添加、删除或编辑基因。使用 EXZACT™,植物研究人员可以测试假设、开发遗传特性并将其引入植物,同时避免传统 DNA 工程工具带来的意外影响。EXZACT™ 加快了特性开发时间并降低了农产品成本,并为作物特性研发建立了新的行业标准。了解更多信息,请访问 www.exzactprecisiontechnology.com。

锌指蛋白:基因组稳定性的守护者
锌指蛋白 (ZNF) 是一类独特而多样的蛋白质,在转录调控、染色质重塑、蛋白质/RNA 稳态和 DNA 修复等基本细胞机制中发挥着关键作用。因此,ZNF 蛋白的错误调节可导致多种人类疾病,从神经发育障碍到多种癌症。考虑到 DNA 损伤修复 (DDR) 抑制在临床上取得了良好的效果,作为同源重组 (HR) 缺陷患者的治疗策略,确定其他潜在的可靶向 DDR 蛋白作为耐药肿瘤细胞中出现的弱点至关重要,尤其是考虑到获得性耐药的负担时。重要的是,越来越多的研究确定了新的 ZNF 并揭示了它们在几种 DDR 通路中的重要性,凸显了它们作为 DDR 抑制疗法新靶点的巨大潜力。尽管如此,仍有许多未表征的含 ZNF 蛋白具有未知的生物学功能。在这篇综述中,我们重点介绍了哺乳动物细胞中 ZNF 蛋白的主要类别和观察到的生物学功能。我们简要介绍了众所周知和新发现的 ZNF,并描述了它们的分子作用以及对人类健康和疾病(尤其是癌症)的贡献。最后,我们讨论了 ZNF 在 DNA 修复机制中的重要性、它们在癌症治疗中的潜力以及利用 ZNF 蛋白作为人类疾病未来治疗靶点的进展。

C2H2锌指蛋白在...中的功能和作用
C2H2 锌指 (C2H2-ZF) 蛋白是人类转录因子的主要类别,它们通过不同的锌指结构域组合实现多种不同的分子功能。肝细胞癌 (HCC) 是最常见的恶性肿瘤之一,也是全球癌症相关死亡的主要原因。越来越多的研究结果支持 C2H2-ZF 蛋白的异常表达在 HCC 的发生和发展中起着重要作用。C2H2-ZF 蛋白参与 HCC 中的各种生物学功能,例如 EMT、干细胞维持、代谢重编程、细胞增殖和生长、细胞凋亡和基因组完整性。抗肿瘤药物耐药性研究也强调了 C2H2-ZF 蛋白在 HCC 生物学功能(EMT、干细胞维持、自噬)和化学耐药性交叉点上的关键作用。最近发现的 C2H2-ZF 蛋白参与调节不同的分子、信号通路和病理生理活动,表明这些蛋白质可能是 HCC 的可能治疗靶点以及诊断或预后生物标志物。

CTCF-DNA复合物的结构,包括所有11个锌指
CCCTC结合因子(CTCF)结合了其11个串联锌(ZF)DNA结合域的哺乳动物ChR量型的增强子和启动子的数十含量。除了12-15 bp的核序列外,某些CTCF结合位点还包含上游和 /或3'下游motifs。在这里,我们分别描述了人类CTCF重叠部分的两个结构,包括ZF1 – ZF7和ZF3 – ZF11与DNA的复合体中的ZF1-ZF7和ZF3 – ZF11,它们将核心序列与3'下游或5'上游基序一起结合在一起。像常规的串联ZF阵列蛋白一样,ZF1 – ZF7 fol-DNA的右手扭曲,每个填充物均占据并识别一个在DNA Major Grove中的三个碱基对的三重态。Zf8 pla ys独特的作用,充当跨DNA或gro的间隔物,并定位ZF9 – ZF11,使其与DNA进行交叉接触。我们将ZF1 – ZF7和ZF8 – ZF11的TW O子分组之间的差异归因于每个纤维内两个位置-6和-5处的残基,而ZF1-ZF7的残基和ZF8 – ZF8 – ZF8 – ZF8-ZF8 – ZF8 – ZF8的ZF1 – ZF7的残基和较小的残基。ZF8也富含碱性氨基酸,该氨基酸使盐桥允许在较小的含量中添加到DNA磷酸盐。较高的特异性Ar ginine-鸟氨酸和谷氨酰胺 - 腺嘌呤相互作用,用于ZFS的常规碱基相互作用位置在常规的碱基相互作用位置上进行补充,也适用于ZF9 – ZF11所采用的跨链相互作用。ZF1 – ZF7和ZF8-ZF11之间的差异可以比例化结构,并且可以促进高实用性CTCF结合位点的识别。

双功能C2H2型锌指转录因子...
ISO avones是由豆类产生的一类二级代谢产物,在人类健康和植物胁迫耐受性中起重要作用。 C2H2锌 - 纤维转录因子(TF)在植物胁迫耐受性中的功能,但对其在大豆(Glycine Max)中的异含量反应中的功能知之甚少。 在这里,我们报告了一个C2H2锌 - 纤维TF基因GMZFP7,该基因调节大豆中的Iso avone积累。 过表达的GMZFP7增加了跨基因根和植物中的ISO avone浓度。 相比之下,沉默的GMZFP7表达显着降低了同avone水平。 代谢组和QRT-PCR分析表明,GMZFP7可以增加苯基丙烷途径的频率。 此外,双 - 荧光酶和电泳动物移动分析测定法表明,GMZFP7通过侵入ISO纤维抗酮合酶2(GMIFS2)(GMIFS2)和3 B-氟酮3 B-羟基酶1(gmff3h1)来调节ISO avone的积累。 在这项研究中,我们证明了GMZFP7通过对竞争性苯基丙烷途径分支的Gateway酶(GMIFS2和GMF3H1)的表达来表达来导致ISO弹药积累,以将代谢流将代谢流引导到ISOOFONE中。 单倍型分析表明,GMZFP7启动子中存在重要的自然变化,P-HAP1和P-HAP3是精英单倍型。 我们的发现提供了有关GMZFP7如何调节苯基丙烷途径并增强大豆ISO avone含量的洞察力。ISO avones是由豆类产生的一类二级代谢产物,在人类健康和植物胁迫耐受性中起重要作用。C2H2锌 - 纤维转录因子(TF)在植物胁迫耐受性中的功能,但对其在大豆(Glycine Max)中的异含量反应中的功能知之甚少。在这里,我们报告了一个C2H2锌 - 纤维TF基因GMZFP7,该基因调节大豆中的Iso avone积累。过表达的GMZFP7增加了跨基因根和植物中的ISO avone浓度。相比之下,沉默的GMZFP7表达显着降低了同avone水平。代谢组和QRT-PCR分析表明,GMZFP7可以增加苯基丙烷途径的频率。此外,双 - 荧光酶和电泳动物移动分析测定法表明,GMZFP7通过侵入ISO纤维抗酮合酶2(GMIFS2)(GMIFS2)和3 B-氟酮3 B-羟基酶1(gmff3h1)来调节ISO avone的积累。在这项研究中,我们证明了GMZFP7通过对竞争性苯基丙烷途径分支的Gateway酶(GMIFS2和GMF3H1)的表达来表达来导致ISO弹药积累,以将代谢流将代谢流引导到ISOOFONE中。单倍型分析表明,GMZFP7启动子中存在重要的自然变化,P-HAP1和P-HAP3是精英单倍型。我们的发现提供了有关GMZFP7如何调节苯基丙烷途径并增强大豆ISO avone含量的洞察力。

锌指蛋白靶向霍乱毒素A(ctxA)基因...
摘要 背景与目的:本研究利用锌指核酸酶(ZFN)技术破坏霍乱毒素基因(ctxA),抑制霍乱弧菌(V. cholera)产生CT毒素。实验方法:设计一个工程化的ZFN,靶向ctxA基因的催化位点,将ZFN编码序列克隆到pKD46、pTZ57R T/A载体和E2-crimson质粒中,转化大肠杆菌(E. coli)Top10和霍乱弧菌,通过菌落计数法评估ZFN的转化效果。结果:转化后的大肠杆菌经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳和蛋白质印迹实验未见表达,ctxA基因测序未见突变,pKD46-ZFN质粒聚合酶链式反应结果为阴性。用含有完整 ZFN 序列的 T/A 载体转化大肠杆菌 Top10 产生 7 个菌落,所有菌落均含有具有自连接载体的细菌。用左阵列 ZFN 转化产生 24 个菌落,其中 6 个含有具有自连接载体的细菌,18 个含有具有载体/左阵列的细菌。用含有完整 ZFN 的 E2-深红色载体转化霍乱弧菌未产生任何菌落。用左阵列载体转化产生 17 个含有具有载体/左阵列的细菌的菌落。使用蛋白质印迹分析捕获左阵列蛋白带。结论和意义:由于缺乏非同源末端连接 (NHEJ) 机制,ZFN 可能脱靶细菌基因组,从而导致致命的双链 DNA 断裂。建议开发针对细菌基因的 ZFN,具有 NHEJ 修复系统的工程包装宿主是必不可少的。关键词:ctxA 基因;基因编辑工具;霍乱弧菌;锌指核酸酶。

与血红蛋白病相关的 C2H2 锌指转录因子
在人类中,β-珠蛋白的特异性畸变会导致镰状细胞病和β-地中海贫血,而这些疾病的症状可以通过增加胎儿珠蛋白 (HbF) 的表达来改善。最近进行的两次 CRISPR-Cas9 筛选以 ~1500 种带注释的序列特异性 DNA 结合蛋白为中心,在表达成人血红蛋白的人类红系细胞中进行,发现了四组 HbF 基因表达的候选调节因子。它们是 (1) 已知可用于控制 HbF 的核小体重塑和去乙酰化酶 (NuRD) 复合蛋白的成员;(2) 七种 C2H2 锌指 (ZF) 蛋白,其中一些 (ZBTB7A 和 BCL11A) 已知可直接沉默成人人类红系细胞中的胎儿 γ-珠蛋白基因;(3) 一些其他不同结构类别的转录因子,它们可能间接影响 HbF 基因表达; (4)DNA 甲基转移酶 1 (DNMT1) 维持 DNA 甲基化标记,这些标记将 MBD2 相关的 NuRD 复合物吸引到 DNA 上,以及相关的组蛋白 H3 赖氨酸 9 甲基化。本文我们简要讨论了这些调节剂(特别是 C2H2 ZF)在诱导 HbF 表达以治疗 β 血红蛋白疾病方面的作用,以及开发安全有效的小分子疗法以调节这种高度保守的血红蛋白转换的最新进展。

提高国产基因组编辑工具“锌指-ND1”的功能性......
修改目标 DNA 的基因组编辑工具是基因和细胞治疗的有力工具。目前主要的基因组编辑工具是CRISPR-Cas,应用最为广泛;其次是TALEN;最后是ZFN,应用最少。其中CRISPR-Cas和TALEN的基本专利将持续到2030年甚至更晚,因此在医疗领域使用需要高额的授权费用。另一方面,ZFN的基本专利已于2020年到期,它是一种可免许可使用的基因组编辑工具。通过将识别DNA的Zinc Finger与切割DNA的FirmCutND1 Nuclease(由广岛大学自主开发)相结合,可以制作出名为“Zinc Finger-ND1”的纯国产基因组编辑工具。然而,构建功能性ZFN并提高其基因组编辑效率极具挑战性。 [研究成果总结] 传统上,创建ZFN的主流方法是从随机重排的ZF中筛选与目标DNA结合的ZF。然而,创建功能性 ZFN 大约需要两个月的时间,这需要大量的时间和精力。另外,人们设计了一种称为“模块化组装”的方法,用于将 ZF 在基因上连接起来,但在制作三指 ZFN(三个 ZF 连接在一起)时,获得功能性 ZFN 的概率约为 5%,由于生产效率低,该方法无法使用。我们假设,手指数量少导致可识别的碱基数量减少,从而导致产生功能性 ZFN 的效率降低。因此,在本研究中,我们采用模块化组装的方式构建了一个6指ZF-ND1(图1),以增加其识别的碱基数量。结果,我们构建的10个ZF-ND1中,有两个被证实具有基因组DNA切割活性,这意味着我们以20%的概率成功获得了功能性ZFN。为了进一步完善ZF-ND1的功能,我们使用结构建模技术(AlphaFold、Rossetta和Coot的分子建模)来模拟ZF和DNA之间的相互作用(图2)。通过与 Zif268(一种与 DNA 结合的天然 3 指 ZF)的 DNA 相互作用模型进行比较,确定了五种候选突变。此外,通过比较与 Zif268 的 DNA 糖磷酸骨架结合的氨基酸,确定了四个突变候选者。当将这九个候选突变逐一引入功能性 ZF-ND1 时,发现其中三个突变(图 3)可提高基因组 DNA 切割活性。 V109K突变使裂解活性提高了5%,并且我们成功在结构建模的基础上增强了ZF-ND1的功能。
锌指抑制剂可抑制免疫检查点分子,提高基因修饰 T 细胞的抗肿瘤活性
在 LV 介导的 ZF-R 递送至 CD3+ 细胞后,MHCI 和 CD5 抑制有效且持久。(A) CD5 基因 mRNA 敲低与 CD5 ZF-R 结合位点 (三角形) 的示意图;颜色越深表示抑制越强。选定的 CD5 ZF-R 以蓝色突出显示。(B) 生成了递送多达两个 ZFR 的 LV 粒子面板,以评估 CD3+ 细胞中的抑制效率。(C) 通过流式细胞术测量 NGFR+/MHCI- 和 NGFR+/CD5- CD3+ 细胞的百分比来量化 CD5 (左) 和 B2M (右) 抑制效率。(D) 通过监测注射到 NXG 小鼠体内 10 周的 NGFR+/MHCI- 和 NGFR+/CD5- CD3+ 细胞来评估 B2M 和 CD5 抑制的持久性。 (E) FACS 图显示注射前(左)和注射后 10 周在血液(中)和脾脏(右)中转导的 CD3+ 细胞中同时出现的 MHCI 和 CD5 抑制。

U2AF26和U2AF35中的锌指域具有不同的功能,包括控制翻译中的作用
抽象最近的工作与剪接体组件U2AF35的两个锌指(ZnF)的点突变与恶性转化有关。然而,令人惊讶的是,对U2AF35 ZNF域的功能知之甚少。在这里,我们分析了哺乳动物U2AF35的ZNF域及其旁系同源物U2AF26的关键功能。两个ZNF都是剪接调节所必需的,而仅ZNF2控制蛋白质稳定性,并有助于与U2AF65的相互作用。这些特征在缺乏ZnF2的U2AF26的自然存在的剪接变体中得到了证实,该变体在激活原代小鼠T细胞时强烈诱导并局部位于细胞质中。在模型T细胞系中使用Ribo-Seq我们为U2AF26在激活基因表达中的细胞质步骤中的作用提供了证据,尤其是翻译。一致地,MS2绑定测定法表明,当定位于模型mRNA的5 rtr时,细胞质U2AF26/35增加了翻译。该法规部分取决于Znf1,因此在核心剪接因子,ZNF域和翻译调节之间提供了联系。总的来说,我们的工作揭示了U2AF26/35及其ZNF领域的意外功能,从而有助于更好地理解其在哺乳动物细胞中的作用和调节。