XiaoMi-AI文件搜索系统
World File Search System同源

CDK12损失驱动前列腺癌的进展,转录复制冲突和旁系同源性CDK13
Jean Ching-Yi Tien, 1,2 Jie Luo, 1,2,14 Yu Chang, 1,2,14 Yuping Zhang, 1,2,14 Yunhui Cheng, 1,2,14 Xiaoju Wang, 1,2 Jianzhang Yang, 3,4 Rahul Mannan, 1,2 Somnath Mahapatra, 1,2 Palak Shah, 1,2 Xiao-Ming Wang, 1,2 Abigail J. Todd, 1,2 Sanjana Eyunni, 1,2 Caleb Cheng, 1 Ryan J. Rebernick, 1,2 Lanbo Xiao, 1,2 Yi Bao, 1,2 James Neiswender, 5 Rachel Brough, 5 Stephen J. Pettitt, 5 Xuhong Cao, 1,2 Stephanie J.), arul@med.umich.edu (A.M.C.)https://doi.org/10.1016/j.xcrm.2024.101758

认知本体论在认知同源性方面:大脑的作用,
图3示意图(改编自Difrisco,Love and Wagner,2020)的“配对gnathostome附属物”。脊椎动物胚胎中肢体芽的发育先于配对附属物的发展(例如鳍和四肢)。双向箭头示意性地描述了组织类型之间的局部激素驱动的相互作用;肢体发育由四个肢体芽中的四个信号中心精心策划。此处包括两个最著名的信号中心,作为带有信号梯度的箭头。ZPA建立了肢体的前后组织组织,AER建立了近端组织。这些信号传导中心形成了一个复杂的,有因果关系的,共同决定肢体身份的必要机制。他们的活动是相互依存的,相互加强(Difrisco,Love and Wagner,2020)。CHIM的上游(其因果“输入”)是相对未分化的,宽松的中胚层胚胎组织; CHIM的下游(其因果“输出”)是具有可识别的组织类型和肢体组织的结构。然后通过进一步的过程来修改这种基本的肢体结构,以确定可能的肢体表型,例如上面讨论的五达乙酰基前肢表型。
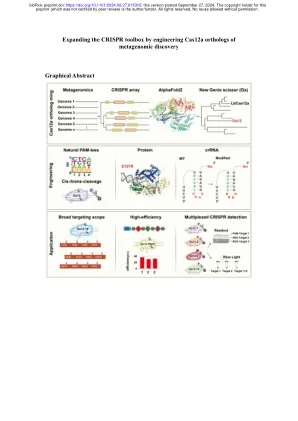
通过工程CAS12A元基因组发现的cas12a直系同源
1农业动物遗传学,育种和繁殖,教育部和猪遗传学和育种部关键实验室,农业和农村事务部,瓦兹洪农业大学,430070 Wuhan,P.R。R.中国。2 Yazhouwan国家实验室(YNL),Sanya Hainan 572025,P。R.China。 3瓦兹胡农业大学的可持续猪生产合作创新中心,430070 Wuhan,P。R.中国。 4 Hubei Hongshan实验室,Huazhong农业大学,Wuhan 430070,P。R.China。 5胰腺疾病实验室,吉安格大学医学院第一家附属医院,杭州310058,P。R.China。 这些作者也同样贡献:Dagang Tao,Bingrong Xu,Sheng Li和Hailong Liu。 *电子邮件:ssxie@mail.hzau.edu.cn; shzhao@mail.hzau.edu.cn; xyli@mail.hzau.edu.cn2 Yazhouwan国家实验室(YNL),Sanya Hainan 572025,P。R.China。3瓦兹胡农业大学的可持续猪生产合作创新中心,430070 Wuhan,P。R.中国。4 Hubei Hongshan实验室,Huazhong农业大学,Wuhan 430070,P。R.China。 5胰腺疾病实验室,吉安格大学医学院第一家附属医院,杭州310058,P。R.China。 这些作者也同样贡献:Dagang Tao,Bingrong Xu,Sheng Li和Hailong Liu。 *电子邮件:ssxie@mail.hzau.edu.cn; shzhao@mail.hzau.edu.cn; xyli@mail.hzau.edu.cn4 Hubei Hongshan实验室,Huazhong农业大学,Wuhan 430070,P。R.China。5胰腺疾病实验室,吉安格大学医学院第一家附属医院,杭州310058,P。R.China。这些作者也同样贡献:Dagang Tao,Bingrong Xu,Sheng Li和Hailong Liu。*电子邮件:ssxie@mail.hzau.edu.cn; shzhao@mail.hzau.edu.cn; xyli@mail.hzau.edu.cn

果蝇 Pten 直系同源基因模型
(A) 果蝇 (Drosophila melanogaster) 和果蝇 (D. miranda) 中 Pten 基因组邻域的同源性比较。细箭头表示果蝇 (D. melanogaster) (上) 和果蝇 (D. miranda) (下) 中目标基因 Pten 所在的 DNA 链。指向右侧的细箭头表示 Pten 在果蝇 (D. miranda) 中位于正 (+) 链上,指向左侧的细箭头表示 Pten 在果蝇 (D. melanogaster) 中位于负 (-) 链上。指向与 Pten 相同方向的宽基因箭头相对于细箭头位于同一链上,而指向 Pten 反方向的宽基因箭头相对于细箭头位于反链上。果蝇 (D. miranda) 中的白色基因箭头表示与果蝇 (D. melanogaster) 中相应基因直系同源,而黑色基因箭头表示非直系同源。灰色箭头表示在两个基因组邻域中都存在但不是同源的基因(在本例中为 Ror),在 D. miranda 中位于 Pten 的上游,但在 D. melanogaster 中位于 Pten 的下游。D. miranda 基因箭头中给出的基因符号表示 D. melanogaster 中的直系同源基因,而基因座标识符是 D. miranda 特有的。(B)GEP UCSC Track Data Hub 中的基因模型(Raney 等人,2014)。D. miranda 中 Pten 的编码区显示在用户提供的轨道(黑色)中;CDS 用粗矩形表示,内含子用细线表示,箭头表示转录方向。后续证据轨迹包括果蝇 (D. melanogaster) 蛋白质的 Spaln(紫色,果蝇 (D. melanogaster) 的 Ref-Seq 蛋白质比对)、NCBI RefSeq 基因的 BLAT 比对(深蓝色,果蝇 (D. miranda) 的 Ref-Seq 基因比对)、TransDecoder 预测的转录本和编码区(深绿色)、成年雌性和成年雄性的 RNA-Seq(分别为红色和浅蓝色;果蝇 (D. miranda) 的 Illumina RNA-Seq 读段比对)以及使用果蝇 (D. miranda) RNA-Seq (SRP009365) 由 regtools 预测的剪接点。所示的剪接点具有最小读取深度 10,其中 10-49、50-99 和 100-499 支持读取分别以蓝色、绿色和粉色表示。 (C) 果蝇 Pten-PB(x 轴)与果蝇直系同源肽(y 轴)的点图。左侧和底部标明氨基酸编号;顶部和右侧标明 CDS 编号,CDS 也以交替颜色突出显示。点图中的间隙表示序列相似性较低的区域。

微同源性介导的末端连接记事
遗传信息的保真度对于细胞功能和生存力至关重要。DNA 双链断裂 (DSB) 对基因组完整性构成重大威胁,需要有效的修复机制。虽然主要的修复策略通常是准确的,但矛盾的是,也存在容易出错的途径。本综述探讨了微同源介导的末端连接 (MMEJ) 的最新进展和我们对它的理解,MMEJ 是一种在生物体中保守的内在致突变 DSB 修复途径。MMEJ 的核心是 DNA 聚合酶 theta (Pol θ ) 的活性,它是一种促进 MMEJ 致突变性的专门聚合酶。我们研究了 MMEJ 活性背后的分子复杂性,并讨论了其在有丝分裂过程中的功能,其中 Pol θ 的活性作为解决持久性 DSB 的最后一搏而出现,尤其是当同源重组受到损害时。我们探索了针对 Pol θ 在癌症治疗和基因组编辑中的有希望的治疗应用。最后,我们讨论了 MMEJ 的进化后果,强调了它在保护基因组完整性和驱动基因组多样性之间的微妙平衡。

浙江同源康医药股份有限公司
本招股说明书的副本连同本招股说明书“附录八 — 交付公司注册处并可供展示的文件”一节所列的文件,已根据《公司(清盘及杂项条文)条例》(香港法例第 32 章)第 342C 条的规定,在香港公司注册处登记。香港证券及期货事务监察委员会及香港公司注册处对本招股说明书或上述任何其他文件的内容概不负责。除非另有公布,否则发售价为每股 H 股 12.10 港元。香港发售股份申请人于申请时须就每股香港发售股份支付发售价12.10港元,并须缴付1.0%经纪佣金、0.0027%证监会交易征费、0.00565%联交所交易费及0.00015%AFRC交易征费。总协调人(以此身份并代表承销商行事)可于我们同意下,于香港公开发售申请截止日期当日早上或之前随时减少全球发售的发售股份数目及╱或发售价。在此情况下,我们将不迟于香港公开发售申请截止日期当日早上在联交所网站www.hkexnews.hk及本公司网站www.tykmedicines.com刊登公告。有关进一步信息,请参阅本招股说明书中的“全球发售结构”和“如何申请香港发售股份”。如果在上市日上午 8:00 之前发生某些事件,总协调人(为其自身及代表香港承销商)有权终止香港承销商根据香港承销协议承担的义务。请参阅本招股说明书中的“承销”。

SIR2抗疾病蛋白的人类同源物通过TLR途径介导免疫
。cc-by-nc-nd 4.0国际许可证。是根据作者/资助者提供的预印本(未经同行评审认证)提供的,他已授予Biorxiv的许可证,以在2024年9月21日发布的此版本中显示此版本的版权持有人。 https://doi.org/10.1101/2024.09.18.613514 doi:Biorxiv Preprint

映射人类基因组中所有旁系同源物的遗传相互作用
此预印本版的版权持有人于2024年7月19日发布。 https://doi.org/10.1101/2024.07.16.603642 doi:Biorxiv Preprint

指向有关人类(同源)细胞材料供应的指南
FDA建议公司为人类细胞材料的收集,存储和运输实施标准程序,尤其是如果在多个地点发生收集的情况下,FDA建议公司实施公司来收集,存储和运输人类细胞材料的标准程序,尤其是在多个地点发生的情况下,在多个地点在符合GMP标准的情况下,没有得出结论。

SRGAP2的人类特异性旁系同源于诱导新苯二醇特征
。cc-by-nc 4.0国际许可(未获得同行评审证明),他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该预印本版本的版权所有者,该版本发布于6月29日,2024年。 https://doi.org/10.1101/2024.06.28.601266 doi:biorxiv Preprint