XiaoMi-AI文件搜索系统
World File Search System增强子

CTCF 拓扑边界的解剖揭示了增强子-致癌基因调控的原理
增强子-基因通讯依赖于拓扑关联域 (TAD) 和由 CCCTC 结合因子 (CTCF) 绝缘子强制执行的边界,但其潜在的结构和机制仍然存在争议。在这里,我们研究了一种通常隔离成纤维细胞生长因子 (FGF) 致癌基因但在胃肠道间质瘤 (GIST) 中被 DNA 高甲基化破坏的边界。该边界包含一系列 CTCF 位点,可强制相邻的 TAD,一个包含 FGF 基因,另一个包含 ANO1 及其推定的增强子,它们在 GIST 及其可能的起源细胞中具有特异性活性。我们表明,边界中四个 CTCF 基序的协调破坏会融合相邻的 TAD,允许 ANO1 增强子接触 FGF3,并导致其强烈诱导。高分辨率微 C 图揭示了 ANO1 增强子和 FGF3 启动子中的转录起始位点之间的特定接触,这种接触与 FGF3 诱导呈定量关系,因此接触频率的适度变化会导致表达的强烈变化,与因果关系一致。

DNA转录的神秘和美丽-Iris Publishers
图9:哺乳动物转录的调节。通过形成染色体环的形成,使DNA的活跃增强子调节区域可与靶基因的启动子DNA区域相互作用。这可以通过RNA聚合酶II(RNAP II)启动Messenger RNA(mRNA)合成,并在基因的转录开始位点与启动子结合。通过锚定在增强子上的一个结构蛋白稳定环,一个固定在启动子上的蛋白质,将这些蛋白固定在启动子上,并连接到形成二聚体(红色锯齿形)。特定的调节转录因子与增强子上的DNA序列基序结合。一般转录因子与启动子结合。当转录因子被信号激活时(此处指示为增强子上的转录因子上的小红色星形所示的磷酸化),增强子被激活,现在可以激活其目标启动子。通过绑定的RNAP IIS在相反的方向上在DNA的每条链上转录活性增强子。介体(一个由相互作用结构中约26个蛋白质组成的复合物)将调节信号从增强子DNA结合的转录因子传达给启动子。

发育生物学学会第83届年会...
摘要书籍程序摘要#1探索阴影增强剂建筑的功能意义Jillian Ness,Yu Wang,Christian Mei,Renata Serio,Renata Serio,Zeba Wunderlich Boston University,美国,美国许多发育基因都受到一套看似冗余的增强剂的调节,这些增强剂可以推动重叠的Spatiotemporpormoral模式。这些多增强器系统被称为阴影增强剂。阴影增强子可以缓冲遗传和环境应力以驱动正常的基因表达模式。使用果蝇胚作为模型,我们表明,前后图案基因的阴影增强子通过结合不同的输入转录因子的不同集来驱动一致的基因表达模式。尽管如此,尚不清楚为什么阴影增强子转录因子结合位点分布在多个增强器上,而不是在单个增强器中。可以想象,阴影增强子使用的机制可以编码为单个增强剂。我们已经生成了增强剂记者,其中消除了阴影增强剂之间的内源性DNA以做出“挤压”配置。令人惊讶的是,我们发现毛压和内源间隔增强子之间的RNA模式,水平和动力学几乎没有差异,这表明阴影功能不需要内源性间距。我们正在构建各个长度的合成污垢增强子,以测试squish增强子保留功能的较低尺寸限制。要理解引起阴影增强子的进化动力学,我们已经使用生物信息学来识别发育阴影增强子集的起源。星期四,我们发现重复事件和可转座元素似乎是果蝇中相对较小的(<10%)的发育阴影增强子出生来源,与我们在小鼠基因组中的发现相比(〜30%)。这些数据表明,合并的单个增强剂可以与不同的阴影增强子相比功能,而不会在压力下损害忠诚度。我们将探测创造和维持阴影增强子的进化动力学,以使其在动物发育中的普遍作用合理化。由NIH授予R01HD095246资助的工作。

快速肌球蛋白超级增强子通过与肌球蛋白基因的竞争性相互作用决定肌纤维表型
成年肌纤维的收缩特性由其肌球蛋白重链异构体含量决定。在这里,我们通过 snATAC-seq 鉴定出重组快速肌球蛋白基因的位点上有一个 42 kb 的超级增强子。通过 4C-seq,我们发现活性快速肌球蛋白启动子通过 DNA 环路与该超级增强子相互作用,导致每个细胞核中单个启动子的激活。包括超级增强子的位点彩虹小鼠转基因模型重现了成年快速肌球蛋白基因的内源性时空表达。通过 CRISPR/Cas9 编辑原位删除超级增强子表明其在控制相关快速肌球蛋白基因方面发挥了重要作用,而删除位点上的两个快速肌球蛋白基因表明启动子对共享超级增强子存在积极竞争。最后,通过破坏快速肌球蛋白的组织,我们发现肢体骨骼肌内的位置异质性,这可能是某些肌病中选择性肌肉易受损伤的原因。

染色质分析确定了对骨骼发育很重要的软骨细胞特异性 Sox9 增强子
转录因子 SRY 相关 HMG 盒 9 (Sox9) 对软骨形成至关重要。SOX9 内部和周围的突变会导致以骨骼畸形为特征的软骨发育不良 (CD)。尽管 Sox9 在此背景下的功能已被充分研究,但调节软骨细胞中 Sox9 表达的机制仍有待阐明。在这里,我们使用全基因组分析来识别位于负责 CD 的近端断点簇中的 2 个 Sox9 增强子。E308(位于 5′ 上游 308 kb)和 E160(位于 5′ 上游 160 kb)的增强子活性与 Sox9 表达水平相关,并且两种增强子在体外均表现出协同作用。虽然小鼠中的单个缺失没有明显影响,但同时缺失 E308 和 E160 会导致侏儒表型,同时软骨细胞中 Sox9 表达减少。此外,在 E308/E160 缺失小鼠中,肢体芽间充质细胞的骨形态发生蛋白 2 依赖性软骨细胞分化严重减弱。最后,我们发现在 E308/E160 缺失小鼠中,Sox9 基因上游的开放染色质区域被重组,以部分补偿 E308 和 E160 的缺失。总之,我们的研究结果揭示了软骨细胞中 Sox9 基因调控的机制,这可能有助于我们理解骨骼疾病的病理生理学。

NSMCE2 是一种新型超增强子调控基因,与乳腺癌预后不良和治疗耐药性有关
在本研究中,我们确定了两个新的超增强子相关基因:NSMCE2 和 MAL2,它们在乳腺肿瘤中高度上调,其高 RNA 水平与乳腺癌患者的不良预后有显著且明确的相关性。为了实现这一目标,我们利用了现有的数据集,其中包含在原发性乳腺肿瘤中确定的超增强子相关基因,以及包含乳腺癌患者的基因表达、基因组和临床结果的公共数据库。通过乳腺癌细胞中的体外药理学超增强子破坏试验,我们证实了超增强子参与了 NSMCE2 和 MAL2 转录本的上调,并通过生物信息学发现高水平的 NSMCE2 与化疗反应不佳密切相关。这在被诊断为侵袭性三阴性和 HER2 阳性肿瘤类型的患者中尤为明显。最后,我们表明,用化疗药物治疗乳腺癌细胞,同时通过超增强子阻断或直接沉默 NSMCE2 基因表达来降低 NSMCE2 基因表达,可以降低细胞活力,从而提高化疗的效果。我们的结果表明,调节新发现的超增强子相关基因 NSMCE2 的转录水平可以改善患者对标准化疗的反应,从而可能改善疾病结果。总之,通过挖掘现有的公共乳腺癌数据集,我们的工作表明,寻找超增强子调节基因及其与患者生存和治疗反应的关联,可能是识别肿瘤特异性(不是经常突变,而是超增强子失调的基因)特征的有效方法。我们的方法为识别预后不良的新生物标志物和改善癌症治疗的潜在药理学靶点提供了一种新途径。

通过染色质相互作用数据的新型归一化方法促进增强子启动器循环的检测
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本版本的版权持有人于2025年1月1日发布。 https://doi.org/10.1101/2024.12.30.630839 doi:Biorxiv Preprint
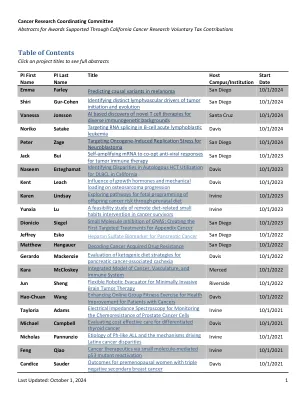
目录
预测黑色素瘤的致病变异 校园:UCSD 首席研究员:Emma Farley 开始日期:2024 年 10 月 1 日 结束日期:2025 年 9 月 30 日 金额:95,000 美元 摘要:癌症的各个方面都由基因表达的变化驱动。增强子是控制特定基因或基因表达的时间、位置和水平的基因组元素,因此,增强子提供基因表达的指令。虽然人们广泛关注蛋白质编码变异和改变蛋白质编码区域的基因组变化,但增强子变异如何促进癌症的发生、进展、转移和对治疗的反应却研究甚少。增强子包含与包括癌症在内的疾病相关的大多数变异,但精确定位致病变异是一项重大挑战,因为它们通常嵌入在大量惰性变异中。我们知识上的这一空白阻碍了充分利用基因组数据的潜力来了解和治疗癌症的努力。清楚了解哪些增强子变异对癌症的各个方面有影响对于了解癌症发生和发展的遗传基础、开发新型疗法、改善诊断和对患者进行分层以进行更有针对性的治疗至关重要。在这项研究中,我们将试行一种新方法来识别导致黑色素瘤基因表达变化的增强子变异。我们是识别增强子中因果变异的专家,这些变异会在发育中的胚胎中改变基因表达和细胞身份。我们发现,低亲和力结合位点对于精确控制基因表达至关重要。增强子中低亲和力位点的普遍使用造成了基因组中的脆弱性,SNV 可以增加增强子中结合位点的亲和力,从而导致功能基因表达获得,从而改变细胞身份。我们已经在心脏和肢体发育的背景下证明了这一点。我们现在希望应用这些知识来深入了解导致癌症各个方面的因果变异。我们计划最初重点关注黑色素瘤的转移、免疫疗法反应和耐药性。在我们的初步研究中,我们已表明,在体细胞黑色素瘤 eQTL 中发现的亲和力优化 SNV 会增加 DAAM1 的表达并增加细胞迁移。成功完成该项目将揭示增强子变体对癌症进展和治疗的贡献。

启动子的全基因组核小体分辨率图
摘要 增强子-启动子环路模型长期以来一直主导着基因调控领域,其中增强子通过物理接触激活其靶基因。然而,由于存在替代机制的证据以及缺乏系统验证(主要是由于缺乏合适的实验技术),该模型的普遍性受到了质疑。在本研究中,我们提出了一种新的基于 MNase 的邻近连接方法,称为 MChIP-C,该方法可以在基因组范围内以单核小体分辨率测量蛋白质介导的染色质相互作用。通过应用 MChIP-C 研究 K562 细胞中以 H3K4me3 启动子为中心的相互作用,我们发现与基于限制性内切酶的 C 方法相比,它具有大大提高的分辨率和灵敏度。这使我们能够将 EP300 组蛋白乙酰转移酶和 SWI/SNF 重塑复合物确定为建立和/或维持增强子-启动子相互作用的潜在候选者。最后,利用已发表的 CRISPRi 筛选数据,我们发现大多数经过功能验证的增强子确实与其同源启动子发生物理相互作用,支持增强子-启动子环路模型。

对基于 CRISPR 的转录激活因子进行系统比较,揭示增强子-启动子相互作用的基因调控特征
基于核酸酶失活 CRISPR/Cas (dCas) 的系统已成为一种强大的技术,可以综合重塑人类表观基因组和基因表达。尽管这些平台的采用越来越多,但它们的相对效力和机制差异尚未完全表征,特别是在人类增强子-启动子对中。在这里,我们系统地比较了最广泛采用的基于 dCas9 的转录激活因子,以及由与人类 CBP 蛋白催化核心融合的 dCas9 组成的激活因子,以及人类增强子-启动子对。我们发现这些平台在不同人类细胞类型中显示出不同的相对表达水平,并且它们的转录激活效率因效应域、效应子募集结构、靶位点和细胞类型而异。我们还表明,每种基于 dCas9 的激活剂都可以诱导增强子 RNA (eRNA) 的产生,并且这种 eRNA 诱导与同源启动子的下游 mRNA 表达呈正相关。此外,我们使用基于 dCas9 的激活剂来证明人类增强子和启动子之间可以存在内在的转录和表观遗传互惠性,并且可以通过将基于 dCas9 的转录激活剂靶向增强子来合成驱动增强子介导的下游启动子的追踪和参与。总之,我们的研究为增强子介导的人类基因表达控制和基于 dCas9 的激活剂的使用提供了新的见解。