XiaoMi-AI文件搜索系统
World File Search System形状

轻巧的运动:印刷液晶弹性体的光反应形状变形
摘要软计算机将需要柔软的材料,这些材料表现出丰富的功能多样性,包括形状变形和光反应。这些功能的组合可以在软计算机中有用的行为,可以通过合成表现出局部响应性的材料来进一步发展。可以通过为直接墨水写作(DIW)制定复合墨水来启用液晶弹性体(LCE)的局部响应(LCE),它们是表现出形状变形的软材料。金纳米棒(Aunrs)可以添加到LCES中,以通过局部表面等离子体共振吸收光后光热形状变化。我们比较了LCE公式,重点是DIW和Aunrs的光响应性打印。不同的三维体系结构的局部响应能力启用了可以振荡,爬网,滚动,运输质量并显示其他独特的致动和运动模式,以响应光线,从而使这些有希望的功能材料用于高级应用程序。

氧气和铁的可用性形状癌细胞的代谢适应
三磷酸腺苷(ATP)输出以及葡萄糖,谷氨酰胺和脂肪酸的利用等之间的糖酵解和氧化磷酸化(OXPHOS)之间的动态变化,导致维持和选择对肿瘤细胞亚基的维持和选择在铁氧化环境中的生长优势。铁在自然界中的三个主要生物化学反应中起重要作用:光合作用,氮固定和氧化呼吸,所有这些都需要参与铁硫蛋白,诸如铁治再蛋白质,细胞色素B,以及复合物I,II,II,III,III,III中的Electron Electon Compranton Chain的Electer链中,这都需要参与铁氧化物硫蛋白。异常的铁硫簇合成过程或缺氧将直接影响线粒体电子转移和线粒体oxphos的功能。更多的研究结果表明,铁代谢,氧利用率和缺氧诱导因子相互调节糖酵解与OXPHOS之间的转移。在本文中,我们进行了综合综述,以提供有关肿瘤细胞中糖性和Oxphos调节的新见解。

设计一个形状的座椅垫以改善姿势(dis ...
摘要 长时间坐着会增加健康问题的风险和不适感。因此,座板设计至关重要,主要受两个因素影响:压力分布和座椅轮廓。对于座板不适,较低的平均压力伴随着较少的不适。此外,接触面积大的座椅轮廓与更舒适相关。因此,我们精确设计(虚拟原型)并实现(物理原型)了一种形状的坐垫,旨在将由于座椅和臀部之间的相互作用而产生的压力分布转化为适合国际人口(包括 P5 女性和 P95 男性)的几何形状。有了这种形状,压力应该更均匀、更低,界面接触面积更大,感知舒适度更高。本文通过可重复且直接的方法描述了虚拟和物理原型设计。此外,还通过与标准平垫进行比较,进行了实验以验证该假设。结果表明,设计目标已经实现:异形垫的压力分布比平垫小,接触面积更大。关键词:产品建模/模型、以用户为中心的设计、设计方法、表面建模、原型制造方法 联系人:Fiorillo,Iolanda 萨勒诺工业工程大学 意大利 ifiorillo@unisa.it

结构,形状,拓扑:分子化学中的纠缠概念
分子结构和分子形状的概念在化学文献中无处不在,在化学文献中,它们通常被视为同义词,在化学教学中不可避免地存在缺点。第三个概念,即分子拓扑,不太频繁,但它是分子研究领域(例如定量结构 - 活性关系)中的参考项。本文提出了对这三个概念的认识论分析,旨在阐明其关系的性质以及它们之间的重点和差异。首先,我们讨论了术语分子结构和分子形状的各种接受。然后,我们研究了这些概念历史上的一些关键里程碑,并从认识论的角度分析了结构,形状和拓扑之间的关系。我们指出了每个概念的区别特征,我们表明它们的语义开放性,在专业的背景下可能是富有成果的,在教学背景下变成了不一致和不准确的来源,这是由于教科书制作的这些术语的误导使用所促进的。最终,我们提出了一个拟合标准,以区分分子形状,分子结构和分子拓扑的概念域。

双重性的双重性和形状距离,对比较神经表示的影响
已经提出了神经网络表示之间的多种(DIS)相似性度量,从而导致了零散的研究景观。这些措施中的大多数属于两个类别之一。首先,诸如线性回归,规范相关分析(CCA)和形状距离之类的措施,都学习神经单位之间的明确映射,以量化相似性,同时考虑预期的不断增长。第二,诸如表示相似性分析(RSA),中心内核比对(CKA)和归一化Bures相似性(NBS)之类的措施都量化了摘要统计数据中的相似性,例如逐个刺激的内核矩阵,它们已经不一致地是预期的。在这里,我们通过观察Riemannian形状距离的余弦(从类别1)等于NB(来自类别2)来统一这两个广泛的方法的步骤。我们探讨了这种联系如何导致形状距离和NB的新解释,并将这些措施的对比与CKA进行对比,这是深度学习文献中的流行相似性度量。

DNA纳米结构引导的蛋白质组装成可编程形状
DNA纳米结构引导的蛋白质将蛋白质组装成可编程的形状Qinyi Lu 1,Yang Xu 2,3,Erik Poppleton 3,Kun Zhou 4,Kun Zhou 4,Petr Sulc 2,3,Petr Sulc 2,3,Nicholas Stephanopoulos 2,3 *亚利桑那州立大学设计与生物仪,亚利桑那州坦佩市,亚利桑那州坦佩85287,美国3分子科学学院,亚利桑那州立大学,亚利桑那州坦佩,亚利桑那州坦佩85287,美国4美国生物医学工程系,乔治亚大学技术和埃默里大学,乔治亚州埃默里大学,乔治亚州3032222222222222. yonggang.ke@emory.edu摘要

不同尖端形状金属鱼叉捕获空间碎片的穿透特性
作者之前研究过使用金属鱼叉清除空间碎片,并开发了相应的数值模拟模型。(6)金属鱼叉尖端的形状被认为对穿透行为有很大影响。但目前主要研究的是锥形鱼叉尖端,其他鱼叉尖端形状的研究还不够。此外,很少有研究考察以斜角方向穿透碎片的情况。如果鱼叉有锥形尖端,当碎片的倾斜角超过鱼叉尖端角的一半时,鱼叉就不会穿透碎片。因此,需要适当设计鱼叉尖端的形状,以便以任何角度穿透碎片。然而,也存在锚太深入或穿过目标的潜在问题,使系绳容易被锚穿透造成的撞击孔的毛边切断。因此,在本研究中,我们研究了不同尖端形状的金属鱼叉与不同穿透条件下的穿透行为之间的关系。

高小班级中的形状和空间委托研究论文
本概述借鉴了 Clements 和 Battista (1992) 的观点,其中添加了 0 级,因为原始模型中存在明显缺陷。此版本而非 van Hiele (1986) 的版本,是因为它最常出现在文献中。虽然“儿童”用于表示对小学教育的普遍期望,但这些级别被认为取决于学习经历而不是年龄。
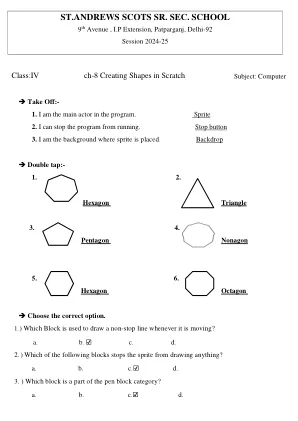
计算机- Ch-8 在 Scratch 中创建形状 (IV) 2024.pdf
填空。1. 多边形是具有 3 条或更多直线和角的二维形状。2. 要计算多边形的旋转度数,请用 360 除以边数。3. 要运行脚本,请单击“执行”按钮。4. 具有六条边的形状称为六边形。

在单个层压轨道的横截面形状上划分的能量沉积过程
在这项研究中,这项研究根据过程条件对熔体池形状变化特性进行了测试分析,以防止传感器在应用定向能源部门工艺技术作为生产嵌入式传感器金属结构的方法时,通过过程的高热能破裂。随着AI技术的发展,结构自我诊断的自我诊断的重要性正在增加,并且随着对结构和传感器融合措施的需求的增加,将传感器插入结构的研究正在扩大。如果将传感器和结构集成到一般制造方法中,则很难避免由过程热能造成的传感器损害。但是,如果您采用激光层压技术,则可以最大程度地减少融合能量以防止传感器破裂。的融合能。本研究比较了通过使用各种激光输出和射线低扫描速度组合的过程条件来比较熔体池组合的熔体池的宽度和深度。目标材料用于SUS316L,激光输出为900〜1,800W,扫描速度定义在800〜1,200mm/min的范围内。根据DIV的分析,随着能量密度的增加,熔体池的宽度增加,并且相同的能量密度证实,熔体池宽度随着产量的增加而增加。中产生的熔体池深度也与能量密度成比例增加,并在1,800W和800 mm/min的过程条件下显示最大深度为700μm。传感器盖的最小厚度,以防止传感器通过在熔体池上方制造。