XiaoMi-AI文件搜索系统
World File Search System逆转录


符合欧洲药典第 2.6.7 章和 USP <77> 草案当前修订版本的新型支原体实时 RT-PCR 检测试剂盒
监管指南要求用于支原体污染检测的 PCR 试剂盒具有高灵敏度,而 DNA 靶标仅在生物体中以低拷贝水平存在,这种灵敏度呈上升趋势。这就是为什么传统的基于 DNA 的 PCR 在试图保持检测的稳健性和可靠性时逐渐达到极限的原因。实时逆转录 PCR 提供了一种克服此问题的智能解决方案。每个在 DNA 水平上可检测到的基因在目标生物体内也可作为转录本。特别是 16S rRNA 区域,一个高度保守的 rRNA 操纵子,是支原体检测的目标,在一个细胞内有多个 RNA 拷贝。RNA 水平上多个靶标的出现有助于用 PCR 检测较少数量的细胞。逆转录聚合酶使 RNA 拷贝可作为 cDNA 靶标,因此与基于 DNA 的基本 PCR 检测相比,可用的 PCR 靶标成倍增加。确实,这种方法无法对 PCR 结果进行任何定量解释,因为 16S rRNA 基因的 RNA 拷贝数非常灵活,但当涉及到需要“是”或“否”答案的质量控制问题时,定量输出不是必需的。这种方法特别简单,因为逆转录已经在 PCR 反应混合物中实施。

载体RNA组(CR001).ai
RBC 的载体 RNA 是合成的 poly-A 核糖核苷酸。正如许多不同的 RT-PCR 系统所证明的那样,用作载体 RNA 的少量 poly-A RNA 不会干扰后续的 RT-PCR,即使使用 oligo-dT 作为逆转录的引物也是如此。

抗病毒逆转录酶的从头基因合成
摘要:细菌使用多种免疫系统来抵御病毒感染,其中许多免疫系统可以感知和靶向外来核酸。防御相关逆转录酶 (DRT) 系统通过利用 DNA 合成为这种免疫策略提供了一个有趣的对照点,但其 DNA 产物的身份和功能仍然很大程度上未知。我们在此展示了 DRT2 系统执行一种前所未有的免疫机制,该机制涉及通过非编码 RNA (ncRNA) 的滚环逆转录进行从头基因合成。对 DRT2 表达细胞中 RT 相关 RNA 和 DNA 配体的无偏分析表明,逆转录通过程序化模板跳跃到 ncRNA 上产生串联的 cDNA 重复序列。然后噬菌体的存在会触发第二链 cDNA 合成,从而产生长双链 DNA。值得注意的是,这种 DNA 产物被高效转录,生成信使 RNA,该 RNA 编码无终止密码子的、永无止境的 ORF(neo),其翻译会导致强大的生长停滞。系统发育分析和对各种 DRT2 同源物的筛选进一步揭示了滚环逆转录和 Neo 蛋白功能的广泛保守性。我们的工作突出了通过 RNA 模板基因创建优雅地扩展了基因组编码潜力,并挑战了沿基因组 DNA 一维轴编码遗传信息的传统范式。

酯液的作用-Midel
ac:资产管理中的关键考虑因素,尤其是在处理老化基础设施时,新解决方案是否可以在不引起干扰或停电的情况下补充现有变电站。我们的天然和合成酯的一个关键好处是,它们可以在高达66 kV的现有矿物油变压器(美洲为69 kV)中进行翻新,包括密封和自由呼吸应用,而无需单位需要任何其他修改,而除了新的垫圈和密封外。可以进行更高的电压变压器;但是,必须对候选资格进行彻底研究。逆转录是一种可靠的解决方案,可以在原位执行,允许网络升级,而无需最终用户产生的资本支出成本。我们有许多例子,在商业场所过夜进行了逆转录,对操作没有影响。

一步三型taqman定量逆转录聚合酶链反应,用于检测猫科罗尼亚病毒,猫Panleukopenia病毒和猫白血病病毒
冠状病毒家族[1]。病毒基因组(约29 kb)包含11个开放式阅读框,它们编码四个结构蛋白和7种非结构性(NS)蛋白质。FCOV根据其致病性分为两种生物型:猫肠病毒(FECV)和猫感染性骨膜炎病毒(FIPV)[2]。FECV感染主要限于肠道,导致轻度,自限制的胃肠道疾病。FIPV会导致致命的多系统,免疫介导的疾病,该疾病是大坝老化的各种组织和器官,腹膜炎甚至死亡是损害的最典型迹象[2,3]。fipv被认为是FECV的突变体,导致病毒致病性和向性欲的变化。然而,可以解释FECV和FIPV的不同致病性的遗传差异仍然不清楚[1,4,5]。根据病毒抗原>
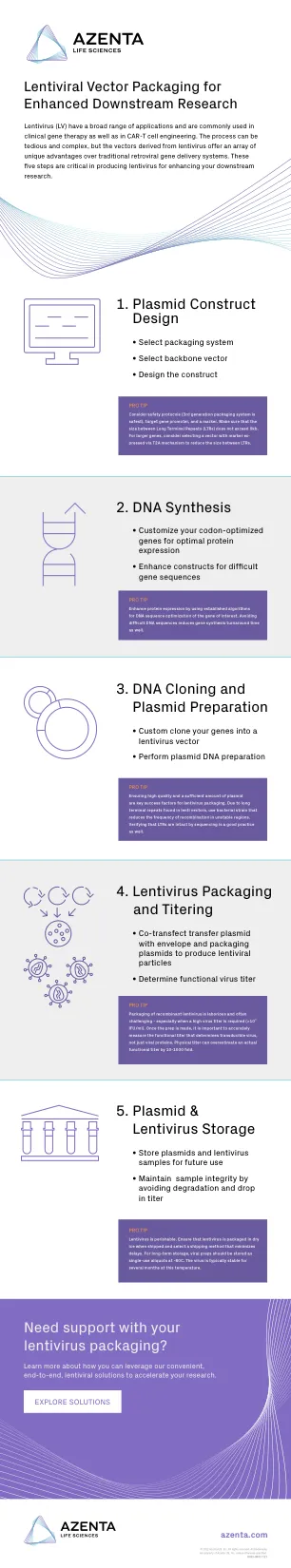
1. 质粒构建体设计 2. DNA 合成 3. ...
慢病毒 (LV) 具有广泛的应用,常用于临床基因治疗以及 CAR-T 细胞工程。该过程可能繁琐而复杂,但慢病毒载体与传统逆转录病毒基因递送系统相比具有一系列独特优势。这五个步骤对于生产慢病毒以增强您的下游研究至关重要。

秀丽隐杆线虫 SEL- 5/ AAK1 调节细胞...
摘要 在秀丽隐杆线虫发育过程中,多个细胞会长距离迁移或伸展突起以到达最终位置和/或获得适当形状。Wnt 信号通路是细胞沿前后体轴迁移或细胞生长的主要协调者之一。Wnt 信号的结果受包括内吞作用在内的各种机制的微调。在本研究中,我们发现 SEL-5(哺乳动物 AP2 相关激酶 AAK1 的秀丽隐杆线虫直系同源物)与逆转录复合物一起在 QL 神经母细胞子细胞迁移过程中作为 EGL-20/Wnt 信号的正调节因子发挥作用。同时,SEL-5 与逆转录复合物的协同作用也是排泄道细胞生长所必需的。重要的是,SEL-5 激酶活性不是其在神经元迁移或排泄细胞生长中发挥作用所必需的,并且这两个过程都不依赖于 DPY-23/AP2M1 磷酸化。我们进一步确定,Wnt 蛋白 CWN-1 和 CWN-2 与 Frizzled 受体 CFZ-2 一起正向调节排泄细胞生长,而 LIN-44/Wnt 和 LIN-17/Frizzled 一起产生抑制其延伸的停止信号。

通过体内产生单链 DNA 进行高通量功能变体筛选
与自然界中存在的巨大变异和基因组工程师设想的巨大变异相比,创建和表征单个遗传变异的规模仍然有限。在这里,我们介绍了逆转录子文库重组 (RLR),这是一种高通量功能筛选方法,其规模和特异性超过了 CRISPR-Cas 方法。我们利用逆转录子的靶向逆转录活性在体内产生单链 DNA (ssDNA),以 > 90% 的效率整合编辑并实现多路复用应用。RLR 同时引入了许多基因组变异,产生了可通过靶向深度测序寻址的汇集和条形码变异库。我们使用 RLR 对合成的抗生素抗性等位基因进行汇集表型分析,展示了相对增长率的定量测量。我们还使用进化细菌的剪切基因组 DNA 进行 RLR,通过实验查询数百万个序列以寻找因果变异,证明 RLR 特别适合利用大量的自然变异库。使用体内产生的 ssDNA 进行汇集实验为探索整个基因组的变异提供了途径。

线虫Panagrellus redivivus
PAT 逆转录转座因子与其他逆转录因子的不同之处在于它们具有“分裂直接重复”结构,即发现内部 300bp 序列重复,每个因子末端约有一半重复。在带有 Northern 印迹的 Panagrellus redivivus 总 RNA 上检测到约 900nt 的非常丰富的转录本,其起始部分映射到 PAT 因子的优先删除部分。潜在对应的 ORF 编码具有羧基末端半胱氨酸基序的 265 个残基的蛋白质,据信这是逆转录因子中 GAG 蛋白的唯一特征。在 Northern 印迹上还检测到一个更暗淡的 1800nt 长的转录本,它位于第一个 ORF 的稍下游。该区域的预测蛋白质序列带有逆转录酶和 RNaseH 的典型基序,如在逆转录因子的 Pol 基因中发现的。肽基序与来自盘基网柄菌的DIRS-1元件最为相似。讨论了使用PAT元件作为秀丽隐杆线虫转座子标记系统的可能性。