XiaoMi-AI文件搜索系统
World File Search SystemAXON

影响耦合轴突束的度量扰动的计算机研究
现在可以通过地球上的仪器探测到引力辐射。与受到人类头骨屏障的电磁辐射相比,引力辐射不受此限制。在 [3] 中,我们通过 MATLAB 模拟展示了引力辐射对人类中枢神经系统中轴突束的影响,这些轴突束之间存在触觉耦合。我们在那里报告说,对于低于 h = 0.09 的应变,对耦合轴突响应的差异时间没有明显影响。考虑到地球接收到的引力波的应变幅度约为 h = 1 e − 21 或更低 [5],我们得出结论,引力波对大脑的信息处理没有影响。然而在本文中,我们得出结论,即使是微弱的引力波也会对轴突束的信息传输产生明确的影响。

髓鞘形成和轴突直径的发育,实现快速而精确的动作电位传导
图 2 蒙古沙鼠梯形体 (TB) 髓鞘的高分辨率图像。抗神经丝相关抗原 (3A10) (a – c) 和神经丝重链 (NFH) (d – f) 的抗体用作轴突标记物。抗髓鞘碱性蛋白 (MBP) 的抗体显示髓鞘。在出生后第 6 天 (a、a')、出生后第 9 天 (b、b 0 ) 和出生后第 13 天 (c、c 0 ) 从 TB 区域沙鼠大脑冠状振动切片中获取共聚焦单光学图像。在出生后第 7 天 (d、d 0 )、出生后第 10 天 (e、e') 和出生后第 14 天 (f、f 0 ) 从 TB 区域沙鼠大脑矢状振动切片中获取共聚焦单光学切片。 (a – f) 轴突标记物 3A10 (a – c) 和 NFH (d – f) 以红色显示,MBP 免疫反应性以绿色表示。(a' – f 0) 相应的 MBP 染色图像。出生后第 6 天 (a、a') 可以看到短的、有时是点状的髓鞘碎片,其间散布着较长的无髓鞘间隙。少突胶质细胞 (白色星号) 积极产生 MBP,用抗 MBP 抗体标记。在出生后第 7 天的矢状切面中,可以看到 TB 纤维的横截面。一小部分 TB 轴突被 MBP 包围,用抗 MBP 抗体标记。出生后第 9 天 (b、b 0),TB 中的大部分轴突都是髓鞘化的。然而,人们可以很容易地注意到一些轴突没有被髓鞘包裹 (白色箭头,b)。在出生后第 10 天的矢状切面上,大多数轴突被髓鞘包裹,但有些没有(白色箭头,e)。到出生后第 13 天(c,c 0)可以看到髓鞘包裹所有轴突。请注意,髓鞘轴突排列非常紧密,以至于很难勾勒出属于单个轴突的髓鞘。在出生后第 14 天的矢状切面上,TB 区域的所有轴突横截面都被髓鞘包裹。比例尺:20 μ m。

皮质乙酰胆碱动力学可通过胆碱能轴突活动和行为状态预测
人们认为乙酰胆碱 (ACh) 在驱动清醒状态下发生的快速、自发的大脑状态转变方面发挥着作用;然而,这些状态变化期间皮质 ACh 活动的时空特性仍不清楚。为了解决这个问题,我们同时对 GRAB-ACh 传感器、表达 GCaMP 的基底前脑轴突和行为进行成像。我们观察到在运动和瞳孔扩张期间轴突和 GRAB-ACh 活动之间存在高度相关性。仅从轴突活动就可以准确预测 GRAB-ACh 荧光,并且局部 ACh 活动在距离轴突较远的地方会降低。对 GRAB-ACh 轨迹进行反卷积使我们能够解释传感器动力学并强调快速清除小 ACh 瞬变。我们训练了一个模型来根据瞳孔大小和跑步速度预测 ACh,

肠道微生物组和健康:机械见解
摘要目的轴突引导途径的失调在胰腺导管腺癌(PDAC)中很常见,但是我们对其生物学相关性的理解受到限制。在这里,我们研究了轴突引导提示SEMA3A在支持PDAC进程中的功能作用。设计,我们整合了人类PDAC的大量和单细胞转录组数据集,以及患者组织的原位杂交分析,以评估PDAC分子亚型中的SEMA3A表达。进行了PDAC细胞系中功能实验的增益和丧失,以剖析SEMA3A如何有助于定义生物学上具有攻击性的表型。导致PDAC组织,SEMA3A由基质元素表达,并选择性地富集在基底/鳞状上皮细胞中。因此,SEMA3A在PDAC细胞中的表达是由基底样表型的细胞中性和细胞脱连的决定因素诱导的。在体外,Sema3a促进细胞迁移以及厌氧性。 在分子水平上,这些表型通过规范SEMA3A-NRP1轴的焦点粘附激酶信号传导增加。 SEMA3A为小鼠PDAC细胞具有更大的转移能力,并有利于肿瘤中与肿瘤相关巨噬细胞的肿瘤内浸润和T细胞密度降低。 从机械上讲,SEMA3A充当巨噬细胞的化学吸引力,并将其极化偏向于M2样表型。 在SEMA3A高肿瘤中,巨噬细胞的耗竭会导致CD8+T细胞的肿瘤内浸润更大,并通过抗肿瘤治疗更好地控制疾病。在体外,Sema3a促进细胞迁移以及厌氧性。在分子水平上,这些表型通过规范SEMA3A-NRP1轴的焦点粘附激酶信号传导增加。SEMA3A为小鼠PDAC细胞具有更大的转移能力,并有利于肿瘤中与肿瘤相关巨噬细胞的肿瘤内浸润和T细胞密度降低。从机械上讲,SEMA3A充当巨噬细胞的化学吸引力,并将其极化偏向于M2样表型。在SEMA3A高肿瘤中,巨噬细胞的耗竭会导致CD8+T细胞的肿瘤内浸润更大,并通过抗肿瘤治疗更好地控制疾病。的结论,我们表明SEMA3A是一个应激敏感的基因座,它通过细胞中的和细胞外在机制促进基底样PDAC的恶性表型。

巨噬细胞耗尽可恢复DRG微环境并防止硼替佐米诱导的神经病中的轴突变性
。cc-by-nc-nd 4.0国际许可证(未经同行评审证明)获得的是作者/资助者,他授予Biorxiv授予Biorxiv的许可,以永久显示预印本。这是该版本的版权持有人,该版本发布于2025年1月24日。 https://doi.org/10.1101/2025.01.22.634362 doi:Biorxiv Preprint

CyFIP2 RNA编辑对肌动蛋白调节,轴突生长和旋转生成的功能影响
Luca La Via 1, Elona Ndoj 1, Matteo Bertoli 1, Veronica Mutti 1, Giulia Carini 1, Alice Filippini 1.2, Federica Bono 1, Chiara Fiorentini 1, Giovanni Ribaudo 1, Alessandra Gianonelli 1, Giuseppe Borsani 1 Isabella Russo 1.2, Alessandro Baron 1.3

Drebrin 调节皮质 II/III 层体感神经元的侧支轴突分支
1. 乔尔·M·多斯坎德 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1 :ROIH 6W %DOWLPRUH 0' -RHOOHGRUVNLQG#JPDLO FRP 'HVLJQHG UHVHDUFK SHUIRUPHG UHVHDUFK FRQWULEXWHG XQSXEOLVKHG UHDJHQWV DQDO\WLF WRROV DQDO\]HG GDWD ZURWH WKH SDSHU 6ULUDP 6XGDUVDQDP 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1 :ROIH 6W %DOWLPRUH 0' VVXGDUV #MKPL HGX 3HUIRUPHG UHVHDUFK DQDO\]HG GDWD 5DQGDO $ +DQG 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1:ROIH 6W %DOWLPRUH 0' 5DQGDO +DQG #JPDLO FRP 'HVLJQHG UHVHDUFK SHUIRUPHG UHVHDUFK FRQWULEXWHG XQSXEOLVKHG UHDJHQWV DQDO\WLF WRROV -DNXE =LDN 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1:ROIH 6W %DOWLPRUH 0' M]LDN #MKPL HGX 3HUIRUPHG UHVHDUFK DQDO\]HG GDWD 0DDPH $PRDK 'DQNZDK 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1 :ROIH 6W %DOWLPRUH 0' PDPRDKGD#ZDNHKHDOWK HGX $QDO\]HG GDWD /XLV *X]PDQ &ODYHO 6XPPHU ,QWHUQVKLS 3URJUDP 1HXUR6,3 6RORPRQ + 6Q\GHU 'HSDUWPHQW RI 1HXURVFLHQFH -RKQV +RSNLQV .DYOL 1HXURVFLHQFH 'LVFRYHU\ ,QVWLWXWH 7KH -RKQV +RSNLQV 6FKRRO RI 0HGLFLQH 1 :ROIH 6W %DOWLPRUH 0' OJX]PDQ #DPKHUVW HGX $QDO\]HG GDWD
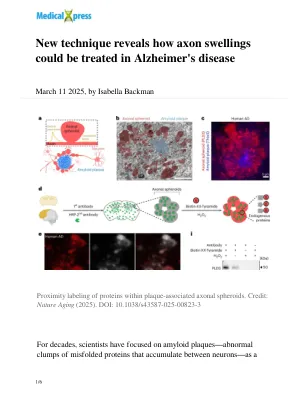
新技术揭示了如何在阿尔茨海默氏病中治疗轴突肿胀
该方法还突出了结构内的多个重要的分子信号通路。其中之一,一种称为MTOR的途径,与细胞生长和代谢有关,在轴突球体中过度活跃。因此,团队进行了一个其他实验,在该实验中,他们将淀粉样蛋白施加到培养皿中的神经元中,导致他们开发类似于人和小鼠组织中轴突球体类似的结构。当他们引入阻断MTOR途径的药理剂时,球体会缩小。这也是在小鼠组织中真实的。

海马engrams生成可变的行为响应和脑范围的网络状态
适当的皮质层压对于认知,学习和记忆至关重要。在体感皮质中,以层状特异性方式详细介绍了锥体式神经元,以决定突触伴侣和整体纤维组织。在这里,我们利用男性和雌性小鼠模型,单细胞标记和成像方法来识别层状特异性侧支的内在调节剂,也称为间隙,轴突分支。我们为II/III层锥体神经元的稳健,稀疏,标记开发了新方法,以获得轴突分支形态的单细胞定量评估。,我们将这些方法与细胞自主的功能丧失(LOF)和过表达(OE)在体内候选筛查中结合在一起,以鉴定皮质神经元轴突分支层压板的调节剂。我们将细胞骨架结合蛋白DREBRIN(DBN1)的作用赋予调节II/III层皮质投射神经元(CPN)侧面轴突在体外的调节中的作用。LOF实验表明,DBN1是抑制II/III层CPN侧支轴突分支在IV层中的伸长的必要条件,在其中,通常不存在轴突通过II/III层CPN分支的轴突分支。相反,DBN1 OE产生过量的短轴突突起,让人联想到未能拉长的新生轴突侧支。结构 - 功能分析暗示DBN1 S142磷酸化和DBN1蛋白结构域已知可介导F-肌动蛋白捆绑和微管(MT)耦合,作为DBN1 OE时侧支分支的必要条件。综上所述,这些结果有助于我们理解调节兴奋性CPN中侧支轴突分支的分子机制,这是新皮层回路形成的关键过程。

内源性钙蛋白酶抑制剂calpastatin减弱了鼠鼠bar综合征的轴突变性
图1内源性HCAST表达可保护急性离体损伤模型中的轴突完整性。三角形(TS)的神经肌肉制剂(WT,n = 3)和Hcast(n = 3)小鼠用抗绞中AB(AGAB)在体内进行内部化研究(A,B),以及(损伤)或没有(对照)正常人(nHs)(nhs)的position(Corsem and Huspect)(CORCE)(CORCE)(cosect)(conterum and)(nhs),以供应(nhs),以供应(nhs),以供介绍。a)WT和Hcast TS运动神经末端(MNT)在37 C下孵育60分钟时,表面AGAB显着降低,而不是0分钟。(b)在用Triton X-100通透性后,在60分钟组中,MNT的总AGAB强度在两种基因型中都恢复正常,这表明AGAB内在化。(c)与对照相比,受伤的WT和Hcast TS的MNT的补体(绿色,E)强度显着增加。(d)神经丝(NF-H,洋红色,E)与对照相比,受伤的WT组织中MNT处的免疫染色强度显着降低,但受到Hcast受伤组织的对照水平的保护。(e)远端神经染色的说明性图像。btx(星号,橙色)和髓磷脂碱性蛋白(MBP,箭头,橙色)分别用于识别Ranvier(Nor)的Mnt和远端节点。虚线的大纲表示没有NF-H染色的位置。比例尺=5μm。 dotplots =平均±S.E.M.在比较治疗效果的数据上进行了未配对的一尾t检验(A,B&C); *表示p <.05。双向方差分析比较治疗(对照与损伤)或基因型(WT vs Hcast),然后对D.的数据进行了Tukey的事后多重比较测试。 * p <.05,***表示p <.001