XiaoMi-AI文件搜索系统
World File Search SystemArtic

研究助理/助理 – ARTIC 测序项目
剑桥大学正在寻找一位积极主动的研究助理或博士后研究员,加入 Ian Goodfellow 教授和 Jenny Molloy 博士的实验室,参与一项由 Wellcome Trust 资助的激动人心的项目,该项目的重点是将基因组监测置于疫情应对的核心。ARTIC 网络是一项全球计划,旨在通过实时基因组测序增强病原体检测和疫情应对。该职位专注于开发和验证传染病监测的测序协议,特别是在资源有限的环境中。研究助理级别的任命取决于拥有博士学位。已提交但尚未获得博士学位的人将被任命为研究助理,一旦获得博士学位,将改为研究助理。

2024年9月供应更新执行摘要
•6月3日,FDA发布了我记得的一堂课,我记得Teleflex/Arrow International的Fiberoptix和Ultraflex Artic Artic气球(IAB)导管套件,分布在2022年5月7日,2022年至2024年4月8日之间。响应5月8日的FDA,向提供者警告提供者持续的安全和质量问题,Getinge正在暂停心脏螺旋桨的促销活动。字母还包括getinge/maquet/datascope Cardiosave混合动力和救援主动脉内气球泵(IABP)设备。Teleflex是美国IAB泵的唯一替代制造商,还为体外膜氧合(ECMO),Fresenius,Abbott,Spectrum,Spectrum和Nautilus/Medtronic的替代制造商致力于跨引用。
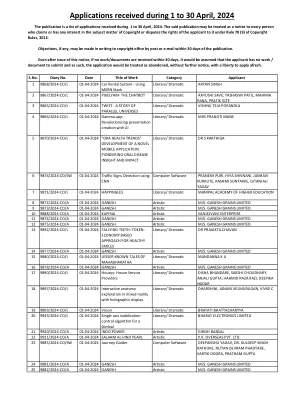
在2024年4月1日至30日收到的申请
85 9914/2024-CO/L 01-04-2024 PAIM-DIT litarary/Dramaatic Omkar Arun Kodulkar,Omkar Santosh Pakhale,Prashant Yuvraj Chavan,Manasi Mahesh Mahesh Lawand 8695/2024-CO/A 01-04-2024 Divine Miracle Artic Logassa Divine Pvt。 div>LTD 87 9894/2024-CO/A 01-04-2024 Divine Dingue登革热杀死艺术Logassa Divine Pvt。 div>LTD 88 9945/2024-CO/L 01-04-2024 @ 60文学/戏剧MR。 Vilas Ravande 89 9899/2024-CO/L 01-04-2024 BHARAT VIBHAJAN KA AITIHASIK

公共咨询 - 美国陆军驻地
北极 Aloha 活动在 PŌHAKULOA 训练中升温为准备第 25 步兵师的联合太平洋多国战备中心 (JPMRC) 和第 11 空降师的北极 Aloha 战备训练演习,基阿库机动区 (KMA) 正在举行多项训练活动。10 月 29 日,驻扎在北卡罗来纳州自由堡的美国空军特种战术小队将进行高空低空开启 (HALO) 跳伞,士兵将进入 KMA。11 月 4 日,车辆和设备将空投到 KMA。此外,美国陆军第 11 空降师将于 10 月 31 日上午进行大规模战术跳伞行动,其中包括数百名伞兵从 KMA 的 C17 上跳下。来自阿拉斯加和夏威夷的部队将一起训练,以提高跳伞能力并加强联合伙伴关系。训练活动将会增加空中交通和噪音。训练活动将在 Saddle Road、Daniel K. Inouye Highway 和 190 号高速公路上进行。我们了解公众有兴趣观看这些活动,但请记住,这些活动都受有利风力条件的影响,可能会被取消或延迟。如果您计划在路边停车,请使用指定的停车区并格外小心。
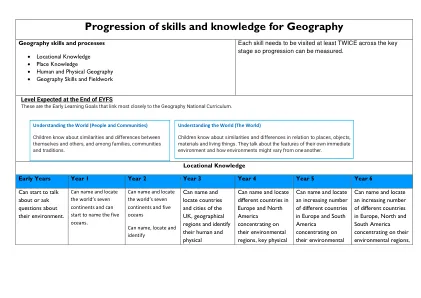
地理技能和知识的发展
biome glacier iceberg Artic Antarctic polar tundra desert monsoon globe map longitude latitude continent ocean Equator North Pole South Pole Northern Hemisphere Southern Hemisphere county Earth solar system universe Tropic of Cancer Tropic of Capricorn Arctic Circle Antarctic Circle Earth's rotation axis compass point N, NE, E, SE, S, SW, W, NW, Mediterranean cliff location resort physical features human特色定居旅游区半岛礁侵蚀沉积潮汐风暴贸易

2024木材产品的碳数据
因此,为了计算表1中的加权平均A4体现碳数据,从EPD确定了每种产品的生产设施。随后通过与各种行业供应商的讨论来设计了该生产设施的现实运输情况(例如HGV的生产设施,原产地,原产地港口到英国港口(例如felixstowe),Defra的平均英国艺术旅行数据)。对于英国和爱尔兰的生产以及通过加来(Ropax)到达的材料,英国的距离是北安普敦的,因为这被认为是英国经济活动的中心。

原始调查|手术机器学习模型用于预测腰椎椎间盘突出症手术后残疾和疼痛的模型
作者分支机构:奥斯陆大都会大学卫生科学学院智能肌肉骨骼健康中心,奥斯陆,挪威(Berg,Gorosito,Fjeld,Fjeld,Haugerud,Haugerud,Storheim,Grotle);挪威奥斯陆奥斯陆大学医院骨科外科司(Berg);奥斯陆大都会大学计算机科学系,挪威奥斯陆(Haugerud Gorosito);挪威奥斯陆奥斯陆大学医院神经病学系(Fjeld);挪威奥斯陆奥斯陆大学医院研究与创新部临床神经科学系(Grotle Storheim);挪威挪威艺术大学临床医学研究所(SOLBERG);挪威北挪威大学医院挪威脊柱外科注册处,挪威,挪威(Solberg)。

纳斯达克 CTA 人工智能指数...
——小型武器、油砂、北极油气勘探和页岩能源被添加到禁止涉足的业务领域,可接受的涉足范围不得超过收入的 5%。热煤发电的可接受涉足范围限制在收入的 5% 以下。热煤发电的容量百分比限制在总容量的 5% 以下。添加了热煤支持产品/服务,可接受的涉足范围不得超过收入的 5%。烟草的可接受涉足范围限制在收入的 5% 以下。除有争议的武器外,所有类别的重大所有权要求均被取消。

第65届年度国际会议-2024
AMI成立于1938年,是该国最古老且著名的科学作品之一。自成立以来,它做出了重要的贡献,可以使Microb iol Ogy的开发人员进行宣传,并在乡下进行研究和教学。目前,该协会有超过5526年的生活和年度成员,约有450名公司成员。该协会在过去64年中发布了季刊《印度微生物学杂志》,每年在该国微生物学的良好中心之一举行全国代表大会。印度微生物学杂志通过出版同行评审到印度和国外研究人员的原始研究结果和研究评论,在世界各地的国家和国际科学研究期刊中都获得了可观的地位。印度微生物学杂志通过出版同行评审到印度和国外研究人员的原始研究结果和研究评论,在世界各地的国家和国际科学研究期刊中都获得了可观的地位。

原始调查|手术机器学习模型用于预测腰椎椎间盘突出症手术后残疾和疼痛的模型
作者分支机构:奥斯陆大都会大学卫生科学学院智能肌肉骨骼健康中心,奥斯陆,挪威(Berg,Gorosito,Fjeld,Fjeld,Haugerud,Haugerud,Storheim,Grotle);挪威奥斯陆奥斯陆大学医院骨科外科司(Berg);奥斯陆大都会大学计算机科学系,挪威奥斯陆(Haugerud Gorosito);挪威奥斯陆奥斯陆大学医院神经病学系(Fjeld);挪威奥斯陆奥斯陆大学医院研究与创新部临床神经科学系(Grotle Storheim);挪威挪威艺术大学临床医学研究所(SOLBERG);挪威北挪威大学医院挪威脊柱外科注册处,挪威,挪威(Solberg)。