XiaoMi-AI文件搜索系统
World File Search SystemFragment

用于治疗阿尔茨海默氏症的BACE1抑制剂药物......
阿尔茨海默病 (AD) 是一种大脑神经退行性疾病,会导致认知功能逐渐受损和记忆力丧失 ( 1 )。AD 主要影响老年患者,全球有超过 5500 万人被诊断患有 AD。目前,尚无治愈方法或改善病情的治疗方法 ( 2 )。到目前为止,大多数获批药物可能有助于控制疾病症状。AD 的标志是神经毒性淀粉样蛋白-β 肽 (Aβ)(主要是 Aβ40 和 Aβ42)在大脑中积累,这被认为是 AD 发病机制的关键特征。在众多解释 AD 发病机制的假说中,20 世纪 90 年代提出的“淀粉样蛋白级联假说”仍然是 AD 药物开发中研究最广泛的概念框架之一 ( 3,4 )。β 位淀粉样蛋白前体蛋白裂解酶 1 (BACE1) 于二十多年前被克隆和鉴定 ( 5,6 )。当时,开发抗淀粉样蛋白-β 药物的前景引起了极大的热情和期待。这种最初的兴奋是可以理解的,因为 BACE1 是第一个启动大脑中 Aβ 产生的蛋白酶,它通过切割淀粉样蛋白前体 (APP) 并形成可溶性 N 端片段和含有膜锚定 C 端片段的淀粉样蛋白-β 肽(图 1)。在接下来的步骤中,γ-分泌酶切割 C 端片段,释放不同长度的神经毒性 Aβ 肽。其中,Aβ42 被认为是 AD 发病和进展的主要原因。从逻辑上讲,这两种蛋白酶都成为抗 AD 的重要药物开发靶点。几个

下一代癌症诊断和治疗
近 50 年前,“第一代”治疗性抗体由鼠源单克隆抗体 (mAb) 组成,目前美国食品药品监督管理局 (FDA) 批准了 30 多种 mAb 用于临床。尽管具有临床潜力,但它们的免疫原性和较大分子量(约 150 kDa)成为其疗效的主要障碍 (1)。这促使人们改进“第二代”,利用抗体片段,例如抗原结合片段 (Fab,约 50kDa) 和单链可变片段 (scFv,约 30kDa);然而,这种方法仍然受到血清半衰期短和聚集诱导的免疫原性的限制 (2)。在骆驼科动物中偶然发现重链抗体 (HcAbs) 引发了最近一波“第三代”抗体浪潮。与传统 mAb 相比,HcAb 仅由两条重链组成,单个可变结构域 (VHH,约 15kDa) 作为抗原结合区。这些纳米级 VHH 被称为“纳米抗体”,分离后可保留完整的抗原结合潜力,使其成为最小的天然抗原结合片段 (3)。纳米抗体促进了商业公司的发展,并已用于生物传感、亲和捕获和蛋白质结晶等应用;然而,它们最显著的潜力在于治疗,尤其是癌症治疗。本综述重点介绍了纳米抗体如何增强各种癌症诊断工具和疗法,包括单独使用和协同作用。最后,本文概述了癌症临床试验中的纳米抗体,分析了障碍,并提出了加快其作为转化癌症疗法实施的潜在策略。


从肉到骨头:法医学的多词方法
质量控制; QQQ,三倍四倍; q-tof,四杆飞行时间; RF,随机森林; RFLP,终末限制片段长度多态性; RMSE,根平方错误; RNA-seq,RNA测序; SBL,结扎测序; SBS,通过合成测序; SCD,心脏猝死; SGD,随机梯度下降; SIDS,婴儿死亡综合症; Silac,氨基酸在细胞培养中稳定的异位标记; Sirm,稳定的同位素分辨代谢组学; SMRT,单分子,实时; SNP,单核苷酸多态性; SQT,简短的QT综合征;德克萨斯州东南部的Stafs应用法医学; STLFR,单管长片段读取; str,短串联重复; SVM,支持向量机; SVM,支持向量机; tadr,胸主动脉

量子达尔文主义、放大和起源......
“从使系统 S 退相干的环境 E 的片段 F 中可以提取多少有关系统 S 的信息?”是量子达尔文主义的核心问题。迄今为止,大多数答案都依赖于 SF 的量子互信息,或通过直接测量 S 提取的数据。这些是真正需要的合理上限,但计算起来要困难得多——片段 F 对于有关 S 的信息的通道容量。我们考虑一个基于不完美 c-not 门的模型,其中可以计算上述所有内容,并讨论其对客观经典现实出现的影响。我们发现所有相关量,例如量子互信息以及通道容量都表现出类似的行为。在与客观经典现实的出现相关的机制中,这包括与不完美 c-not 门的质量或 E 的大小无关的缩放,甚至几乎与 S 的初始状态无关。

机械型 - repisalud
补充图8。TEV蛋白酶的HALOTAG-TEV-TITIN消化的代表动力学。(a)用TEV蛋白酶消化PSOAS纤维。消化在不同时间点停止了通过在Laemmli缓冲液中煮沸的等分试样,并通过1.8%SDS-PAGE和COOMASSIE染色分析结果。对应于云蛋白的条带,并指出完整和裂解的钛分子。(b,c)通过光密度测定法定量钛带。nebulin强度用于标准化(黑色,完整的滴定;蓝色圆圈,A频段滴定片段;红色三角形,I频段滴定片段)。实线是指数拟合,表示速率常数。这些数据表明,在30分钟的反应时间,在此特定实验中,Halotag-Tev Titin的消化> 99.9%(n = 1实验分析每个消化时间的一个样本,在6个实验中获得了类似的结果)。
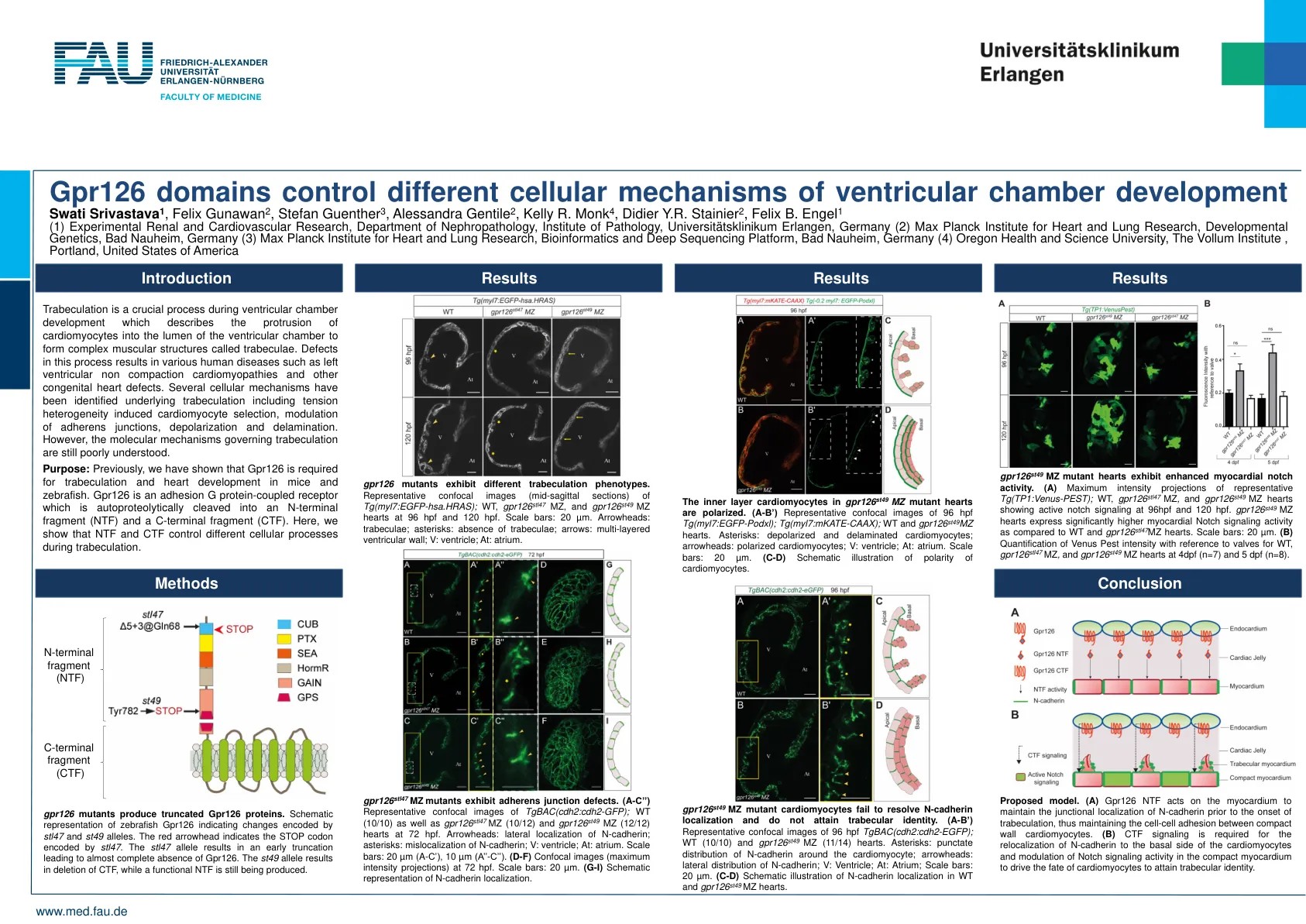
Gpr126 结构域控制心室发育的不同细胞机制 Swati Srivastava 1、Felix Gunawan 2、Stefan Guenther 3、Ale
小梁形成是心室发育过程中的一个关键过程,它描述了心肌细胞突出到心室腔内形成称为小梁的复杂肌肉结构。该过程中的缺陷会导致各种人类疾病,例如左心室非致密化性心肌病和其他先天性心脏缺陷。已经确定了小梁形成的几种细胞机制,包括张力异质性诱导的心肌细胞选择、粘附连接的调节、去极化和分层。然而,控制小梁形成的分子机制仍然不太清楚。目的:之前,我们已经证明 Gpr126 是小鼠和斑马鱼小梁形成和心脏发育所必需的。Gpr126 是一种粘附 G 蛋白偶联受体,可自蛋白酶切为 N 端片段 (NTF) 和 C 端片段 (CTF)。在这里,我们表明 NTF 和 CTF 在小梁形成过程中控制不同的细胞过程。

DNA取证模块No.21:DNA测序
3.1 Maxam – Gilbert测序(化学裂解)方法1976- 1977年,Allan Maxam和Walter Gilbert建立了一种基于DNA的化学改变和在精确基础上的裂解的DNA测序技术。该技术需要放射性标记向一端,并纯化要测序的DNA片段。化学处理形式在单个四个反应(G,A+G,C,C+T)中以四个核苷酸碱基中的一两个或两个的一小部分破裂。因此,从放射性标记的末端到每个分子的第一个“切割”位点产生了一系列标记的残留序列。这四个反应的片段在凝胶电泳中相互组织,以通过尺寸隔离为大小。要观察碎片,将凝胶暴露于X射线膜上以进行放射自显影,根据放射性标记的DNA片段形成一系列暗带,可以从中推断出该排列。

Nebnext Ultra II DNA库Prep套件,用于Illumina,用于Illumina(独特的双索引UMI适配器DNA)E7645 E7645 E7103手动
在人DNA中,有4-6%的胞嘧啶是甲基化的,其中60-90%的甲基化胞嘧啶在CpG位点(1,2)。相比之下,微生物物种中CpG位点的甲基化很少见。NEBNEXT微生物组DNA富集试剂盒使用一种简单而快速的基于磁珠的方法来选择性结合和去除CpG-甲基化的宿主DNA。该方法使用MBD2-FC蛋白,该蛋白由甲基化的CpG特异性结合蛋白MBD2组成,该蛋白与人IgG的FC片段融合在一起。FC片段很容易与蛋白A结合,从而有效地附着在蛋白A结合的磁珠上。该蛋白质的MBD2结构域特异性结合与CpG甲基化DNA。磁场的应用然后拔出CpG-甲基化(真核)DNA,将非CPG-甲基化(微生物)DNA留在上清液中(3)。如果需要,可以洗脱在磁性珠粒中捕获的宿主DNA,并为此提供协议。

蛋白酶抑制剂诱导番茄叶中的诱导因子活性位于从细胞壁酶释放的寡糖中
通过攻击害虫或其他机械损伤释放出一种假定的伤口激素,该激素在整个植物中释放出诱导叶子以引发叶子来引发合成并积聚两个丝氨酸内肽酶的蛋白质含量(1)。该蛋白酶抑制剂诱导因子(PIIF)一直与大小变化的多糖始终相关(2),这表明PIIF活性可能与特定的糖序或结构固有。最近,MR 5000- 10,000的高活性番茄PIIF部分被证明是果多糖。它的位置类似于酶促产生的nicamore细胞壁的碎片,该薄膜壁是200,000的MR,其具有与番茄PIIF相似的效率(3)。该证据表明PIIF活性可能与植物细胞壁的结构成分有关。但是,鉴于大小的大小。番茄果果多糖和nicamore细胞壁碎片均可质疑它们在体内受伤后是否会通过植物血管系统迅速运输。- 在这种交流中,我们报告了一种纯galactu -ronase纯化。真菌根瘤菌(4)将番茄piif降解为寡糖,当蛋白酶抑制剂I的活性诱导剂提供给切除的番茄叶时。我们还表明,部分纯化的两个末代乳乳糖酶的混合物。番茄水果,将番茄PIIF和纯化的番茄细胞壁降解为PIIF活性寡糖。这些结果表明,细胞损伤在体内产生的PIIF活性位于植物细胞壁的小水解碎片中。