XiaoMi-AI文件搜索系统
World File Search SystemGF

2024 年中期报告
完善组织架构,体现业务重点。建筑技术业务整合至 GF Building Flow Solutions(原 GF Uponor),而原 Uponor Infrastructure 业务则并入 GF Piping Systems 的公共设施业务

2024 年处方集 - 涵盖的处方药清单
CalChoice Bronze HDHP NG 1, CalChoice Bronze HDHP NG 3, CalChoice Bronze HMO NG 2, CalChoice Gold HMO NG 2, CalChoice Gold HMO 3, CalChoice Platinum HMO NG Platinum 2, CalChoice Platinum 3, CalChoice Silver 1, CalChoice Silver 2, CalChoice Silver 3, Gold 1, Gold 2, Gold 3, Gold 4, Gold 5, Gold 6, Gold 7, HDHP , HDHP NG 2 L, HDHP NG 3 L, HDHP NG 4 L, HDHP NG 5 L, HMO GF 1, HMO GF 2, HMO GF 4, HMO GF 5, HMO GF 6, HMO GF 7, HMO NG 10 L, HMO NG 11 L, HMO NG 12 L, HMO GF 13 L, HMO.NG 14 L, 15 L, 16 L, 17 L, 18 L, 19 L, 20 L, 21 L, 22 L, 23 L, 24 L, 25 L, 26 L, 27 L, HMO 28 L, 29 L, 30 L, 31 L, 32 L, 33 L, 34 L, 35 L, 36 L, 37 L, 38 L, 39 L, 40 L, 41. L, 42 L, 43 L, 44 L, 5 L, 6 L, 7 L, 8 L, 9 L, Platinum 1, Platinum 2, Platinum 3, Platinum 4, Platinum 7, Platinum 8, POS NG 3 L, Sharp HealthCare HMO NG 1 L (Premium), Silver HMO 1, Silver HMO 2, Sharp HealthCare HMO 2 L (Basic), HMO GF 1 L, HMO GF 3 L, HMO GF 4 L, HMO GF 6 L, HMO GF 14 L, HMO GF 15 L, POS NG 9 L, POS NG 10 L, POS NG 11 L, POS NG 12 L, POS NG 13 L, POS NG 14 L, POS NG 15 L, POS NG 16 L, POS NG 17 L, POS NG 18 L, POS NG 19 L, POS NG 20 L, HDHP POS 21 L, HDHP POS 22 L, HDHP P OS 23 L, Platinum POS NG1, Gold POS NG1, Silver POS NG1, Custom Employer Groups

一种面向运动想象EEG 信号处理的高效脑机接口芯片
∗ 基金项目 : 国家自然科学基金 (61072135,81971702), 中央高校基本科研业务费专项 (2042017gf0075,2042019gf00720), 湖北

哥德尔不完备定理及其对计算的启示
哥德尔的两个不完备性定理中的第一个指出“任何一致的形式系统 F,只要其中可以执行一定数量的基本算术,都是不完备的”6。这意味着 F 中存在既不能证明也不能反驳的陈述(F 中的“哥德尔句”可以称为 GF)。每个系统都有自己的哥德尔句。虽然可以定义一个新的、“更具包容性”的系统 F',并由整个 F 以及之前的哥德尔句 GF 作为公理组成,但这不会产生一个现在完整的系统,因为该定理也适用于修改后的 F 版本,因此 F' 也不完整。因此,GF 将成为 F' 中的一个定理,这并不与哥德尔第一定理相矛盾,因为 GF 在 F 中无法证明,而不是在 F' 中。然而,由于第一定理适用于F',因此存在一个新的哥德尔句子GF',证明F'也是不完整的。

使用 12LP 和 12LP+ 的 AI 云加速器
GlobalFoundries® (GF®) 的 12LP 和 12LP+ AI 加速器解决方案可以帮助解决这些内存和功率瓶颈问题,同时加速 AI 应用。这两款基于 FinFET 的解决方案提供 1 GHz 以上的性能,并采用专用 AI 创新,可显著提高功率效率和面积优势。12LP+ 以 GF 成熟的 14LPP/12LP FinFET 解决方案为基础,GF 已出货超过一百万片此类晶圆。
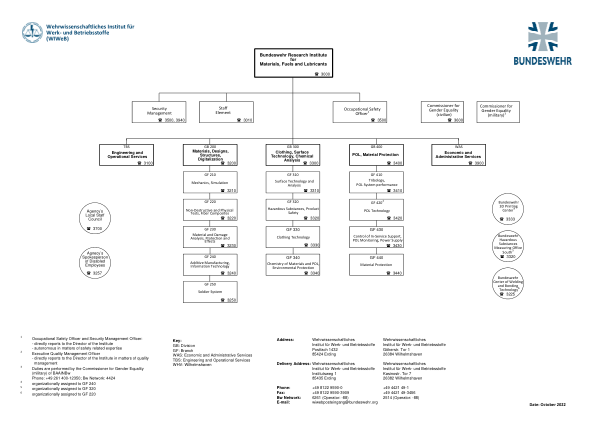
军事材料与操作材料科学研究所 (WIWeB)
1 职业安全官和安全管理官: - 直接向研究所所长汇报 - 在安全相关专业知识方面具有自主权 2 执行质量管理官 - 在质量管理事务上直接向研究所所长汇报 3 职责由 BAAINBw 的性别平等专员(军事)履行 电话:+49 261 400-12350;Bw 网络:4424 4 组织上分配给 GF 240 5 组织上分配给 GF 320 6 组织上分配给 GF 220

新闻稿:欧博诺交易即将完成
为了加速实施 GF 管路系统 2025 战略并进一步受益于全球大趋势,GF 将启动一项雄心勃勃的价值创造计划,以充分发挥此次收购的潜力。GF 管路系统将专注于工业和公用事业业务,为客户提供可持续的无泄漏管道解决方案,用于微电子、水处理、能源、化学加工、海洋、数据中心或水和天然气公用事业等众多终端市场。Uponor 的基础设施业务将成为 GF 管路系统的一部分,并将进一步加强该部门的产品组合,为客户提供综合解决方案。在新部门 GF Uponor 中,建筑技术业务将合并,提供安全清洁的水以及节能供暖和制冷的互补产品组合。互补业务的整合为两个部门开发具有高潜力的市场提供了理想的平台。由于 Uponor 品牌代表着高品质和创新,它将推动智能建筑流量解决方案的增长。

力量到达和X的力量 - 开发新存储概念的历史和结果微生物
摘要:目前,中国的“浓缩物”,“浓缩物 +干草”和TMR“总混合口粮”进食模式的犊牛通常尚不清楚实际生产中的三种分子调节机制。这项研究旨在探索中国荷斯坦犊牛最合适的喂养模式,以改善瘤胃发酵功能和犊牛的生长性能。在这方面,研究了瘤胃微生物与宿主代谢之间的相互作用。GF组的瘤胃体积和犊牛的重量显着高于GFF和TMR组中的瘤犊牛(P <0.05),而GF组的犊牛瘤pH值为6.47〜6.79。宏基因组学分析表明,GF和GFF犊牛的瘤胃微生物组的相对丰度较高,甲烷二磷,甲烷磷和甲诺氏菌具有较高的相对丰度(p <0.05)。prevotella多含糖果在GF犊牛的瘤胃中(p <0.05)的含量更高,这表明GF组犊牛具有更强的发酵糖的能力。值得注意的是,与TMR组相比,在丙酮酸代谢途径中,在GF犊牛中显着上调了磷酸烯醇丙酮酸羧化酶,并且丙酮酸磷酸二酮酶显着下调。代谢组学结果表明,在GF犊牛中,Ursodoxycholic的上调显着上调,并且大多数差异代谢产物都富含胆汁分泌途径。协会分析研究发现,Prevotella和Ruminococcaceae的微生物可能与宿主合作,这有助于消化和吸收脂质,并使犊牛的生长更好。这三种喂养模式具有相似的效果,但是“ GF”喂养模式对有关瘤胃形态,含量生理学和微生物的个人生长和瘤胃发展更为有益。此外,瘤胃微生物和宿主的协同作用可以更有效地水解脂质物质并促进脂质的吸收,这对犊牛的生长具有很大的意义。


文章/书籍信息 - T2R2
我们提出了一种方案,通过量子计算机上的统计抽样来构建相互作用电子系统的单粒子格林函数 (GF)。尽管电子自旋轨道的产生和湮灭算符的非幺正性使我们无法有选择地准备特定状态,但已证明量子比特可以进行概率状态准备。我们提供配备最多两个辅助量子比特的量子电路,以获得 GF 的所有组件。我们基于幺正耦合簇 (UCC) 方法对 LiH 和 H 2 O 分子的 GF 构建进行了模拟,通过比较 UCC 方法中的准粒子和卫星光谱以及全配置相互作用计算的光谱来证明我们方案的有效性。我们还通过利用 Galitskii-Migdal 公式来检查采样方法的准确性,该公式仅从 GF 中给出总能量。