XiaoMi-AI文件搜索系统
World File Search SystemGenome
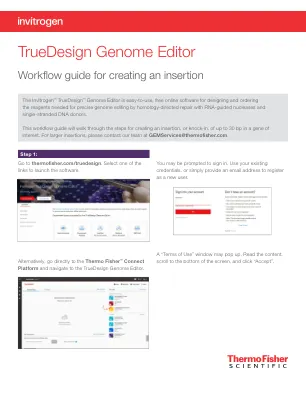
TrueDesign 基因组编辑器
要查看 TALEN 靶标,请单击表格中的 TALEN 选项卡,每个 TALEN 对都会显示类似的信息。当敲入位点 10 bp 范围内没有 PAM 位点,或者 gRNA 的效率和特异性不理想时,建议使用 TAL 效应核酸酶 (TALEN) 对。设计结果表中的绿色复选标记将指示推荐的技术。有关 TALEN 技术的更多信息,请访问 thermofisher.com/tal 。

基因组项目–Write
负责任的创新基因组合成是基因工程工具的逻辑扩展,该工具已在生物技术行业中安全使用了约40年,并提供了重要的益处。然而,最近的技术进步 - 例如,标准化的基因部分,全基因组合成和群集定期散布的短篇小说(CRISPR) - CAS9基因组编辑技术(3,4) - 彻底改变领域(5)。某些申请是有争议的;特别是人类种系编辑引发了激烈的道德辩论(6)。随着人类基因组规模的合成似乎越来越可行,这是及时的统治,讨论和应用大型基因组工程技术的协调科学努力。hgp-写作将需要从一开始就需要公众参与和考虑道德,法律和社会信息(ELSI)。负责任的创新不仅需要ELSI,还涉及通过及时,详细的咨询来确定对科学家和更广泛的公众的共同目标。

基因组编辑工具
瑞典皇家科学院决定将 2020 年诺贝尔化学奖授予 Emmanuelle Charpentier 和 Jennifer A. Doudna,以表彰他们开发了一种基因组编辑方法。引言 1953 年,JD Watson 和 FHC Crick 报告了 DNA 的分子结构 [1]。从那时起,科学家们就一直试图开发能够操纵细胞和生物遗传物质的技术。随着 RNA 引导的 CRISPR-Cas9 系统的发现,一种简单有效的基因组工程方法现已成为现实。这项技术的发展使科学家能够修改各种细胞和生物中的 DNA 序列。基因组操作不再是实验的瓶颈。如今,CRISPR-Cas9 技术被广泛应用于基础科学、生物技术和未来治疗学的开发 [2]。在原核生物中发现 CRISPR-Cas 系统。最终导致发现用于基因组编辑的强大的 CRISPR-Cas9 系统的工作始于鉴定细菌和古菌中存在的重复基因组结构。 1987 年,一份报告指出大肠杆菌基因组中存在一个不寻常的重复结构,该结构包含五个高度同源的 29 个碱基对 (bp) 序列,包括 14 bp 的二元对称序列,其间散布着 32 bp 的可变间隔序列 [3]。几年后,在嗜盐古菌 Haloferax mediterranei 的基因组中也发现了类似的重复结构,其中有 14 个几乎完全保守的 30 bp 序列,以规律的距离重复 [4]。后续的生物信息学分析表明,这些类型的重复在原核生物中很常见,且都具有相同的特殊特征:一个短的部分回文元素成簇出现,并被独特的恒定长度的中间序列隔开,这表明其起源于祖先并具有高度的生物学相关性 [5]。从此引入了术语 CRISPR,这是成簇的规律间隔的短回文重复序列的缩写 [6]。了解 CRISPR 功能的重要一步是鉴定出 CRISPR 相关 (cas) 基因,这是一组仅存在于含有 CRISPR 的原核生物中且始终位于 CRISPR 相邻位置的基因。鉴定出的 cas 基因编码的蛋白质具有解旋酶和核酸酶基序,表明其在 DNA 代谢或基因表达中发挥作用 [6]。与 CRISPR 的关联被用作定义特征,在接下来的几年中,描述了许多 Cas 蛋白亚家族 [7, 8]。CRISPR 位点的功能重要性一直难以捉摸,直到 2005 年,研究人员注意到独特的 CRISPR 序列来自可传播的遗传元件,例如噬菌体和质粒 [9-11]。携带这些特定序列的原核生物似乎可以免受感染,因为含有与间隔序列匹配的序列(称为原间隔序列)的质粒或病毒通常不存在于携带间隔序列的原核生物中 [9, 11]。这些相关发现表明 CRISPR 在原核生物防御入侵外来 DNA 方面发挥着作用,间隔序列被描述为“过去‘遗传攻击’的记忆” [10]。已经证明 CRISPR 转录成长 RNA

基因组编辑漫画
资料来源: Fridovich-Keil, JL (2023 年 12 月 8 日)。基因编辑。大英百科全书。https://www.britannica.com/science/gene-editing • 基因组编辑是如何使用的? (2017)。国家人类基因组研究所。https://www.genome.gov/about-genomics/policy-issues/Genome-Editing/How-genome-editing-is-used • 什么是基因组编辑和 CRISPR-Cas9? (2022 年 3 月 22 日)。MedlinePlus。https://medlineplus.gov/genetics/understanding/genomicresearch/genomeediting/

秘鲁基因组计划
Heinner 1,2,3, Omar Caceres 1, Cesar Sanchez 1, Carlos Padilla 1, Omar Trujillo 1, Victor Border 4,5, Luis Jaramillo-Valverde 2, Julio A. Carolina Silva-Carvalho 6, Mary Horton 7, Cristina M Lanata 7, Alessandra Carnevale 8, Sandra Romero-Hidalgo 8, Victor Acuña-class 10, Pedro Novoa-Bellota 11, Roberto Frisancho 12, Ruth Shady-Solis 11, Pedro Flores-Villanueva 2, Timothy D. O'Connor 4, Manuel Corpas 13,14,15, Eduardo Tarazona-Santos 6 1 National Institute of Salud, Lima, Peru 2 Inbiomedic Research and Technology, Lima From Huanuco, Huanuco, Peru 4 University of Maryland - Institute for Health Computing, 5 Institute for Genomes Sciences, University of Maryland School of Medicine 6 Department of Genetics, Ecology and Evolution, Institute of Biological Sciences, Federal University of Minas Gerais, Belo Horizonte, Brazil 7 National Human Human Genome Research Institute National Genomic Medicine (Inmegen), Mexico City, Mexico. 9 墨西哥墨西哥城 INMEGEN 化学学院生物系应用人群基因组学中心 10 墨西哥墨西哥城国家人类学和历史学院 (ENAH) 11 秘鲁文化部 003 执行单位卡拉尔考古区,秘鲁利马

纳米封装的 CRISPR 基因组
基于 CRISPR 的基因组编辑技术的出现彻底改变了分子生物学,为操纵遗传物质提供了无与伦比的精度。这项变革性技术为医学、农业和生物技术的突破铺平了道路。然而,CRISPR 的全部潜力仍然受到一个关键瓶颈的限制:将其成分有效地递送到靶细胞。脱靶效应、免疫原性和有限的组织特异性等挑战继续阻碍其从实验室到临床的转化。1 为了克服这些障碍,研究人员现在正在探索纳米封装作为优化 CRISPR 递送的下一代策略。2 CRISPR 递送传统上依赖于病毒载体、化学转染(脂质转染)和物理方法,如电穿孔/核转染和微注射。3 虽然病毒载体效率很高,但它们的使用受到免疫原性、尺寸限制和插入诱变风险的限制。脂质转染使用


基因组报告示例
基因组坐标位置渗透载体表型基因覆盖g.8003996666delc(Chr17,grch38)外显子3高3个高note note note note note note note note 15倍变体解释:p.ala83valfsx84在CCDC40中的p.ala83valfsx84变异,先前在19个雄性和7型杂质的helel helesozygous and pc n hymozygous and pc contia和7 compio and syzygous and;在1个纯合受影响的亲戚中与疾病隔离(Becker-Heck 2011 PMID:21131974,Nakhleh 2012 PMID:22499950,Antony 2013 PMID:23255504,Zariwala,2013 PMID:23891469)。该变体已在gnomad(http://gnomad.broadinstitute.org)中鉴定出0.074%(860/1167354)的非欧洲欧洲染色体。但是,此频率足够低,可以与隐性等位基因频率保持一致。在Clinvar中也报道了这种变体(变体ID 31069)。该变体被预测会引起移架,从而改变蛋白质的氨基酸序列,从位置83开始,并导致下游的过早终止密码子84氨基酸。然后预测这种改变会导致截短或不存在的蛋白质。功能研究表明,CCDC40功能的丧失导致纤毛结构和运动异常(Becker-Heck 2011 PMID:21131974)。总而言之,该变体符合标准,该标准被归类为常染色体隐性原发性睫状运动障碍的致病性。ACMG/AMP标准应用:PVS1,PM3_VERYSTRONG,PM2_SUPPORTING,PP1。疾病信息:原发性睫状运动障碍是一种罕见的遗传病,在遗传上是异质的。它与复发性呼吸道感染,内脏异常定位以及不育有关。这是由于器官和组织衬里发现的纤毛和鞭毛的运动性异常。呼吸道感染,粘液清除率降低,鼻塞和慢性咳嗽始于幼儿,可能导致支气管扩张。Situs Inversus Totalis是所有内脏器官的镜像逆转,在40-50%的个体中发现。雌性运动障碍的雄性由于精子运动异常而经常是不育的,而患有这种疾病的女性有时可能是由于输卵管中的纤毛异常引起的。其他症状可能包括大脑中的复发性耳朵感染和脑积水。Pathogenic variants in CCDC40 contribute to 3-4% of primary ciliary dyskinesia (Medline Plus: https://medlineplus.gov/genetics/condition/primary-ciliary-dyskinesia, GeneReviews: https://www.ncbi.nlm.nih.gov/books/NBK1122).家族性和生殖风险疾病患病率(估计)载体频率(估计)生殖风险(估计)1/16000(https://medlineplus.gov/genetics/conditics/condition/primary-ciliary-ciliary-ciliary-dyskinesia)

脉冲中的基因组编辑
限制脉冲潜在产量的主要限制因素包括除了社会经济因素以外的脉冲生长区域中普遍存在的生物和非生物应力。在生物胁迫中,与根腐病配合物相结合的镰刀菌可能是最广泛的疾病,除了干根腐烂和锁骨腐烂外,还会造成鹰嘴豆的巨大损失。虽然镰刀菌,无菌性摩西和植物疫病会导致鸽子,黄色马赛克,尾虫叶斑,粉状霉菌和叶片皱纹和叶片造成大量损失,并在Vigna作物(Mungbean和Urdbean)中造成了相当大的损害。在鹰嘴豆和鸽子中的革兰氏荚虫(Helicoverpa Armigera)中,岩豆和鸽子中的革兰氏pod虫,木豆中的豆荚在乌尔德比恩和蒙比e造成严重损害各自的作物的豆荚,粉丝,粉丝,jassids和thrips。bruchids是储存的脉冲晶粒中最严重的害虫,在管理中需要最高优先级。杂草也会大大损失脉冲。最近,线虫已成为许多地区成功种植脉冲的潜在威胁。
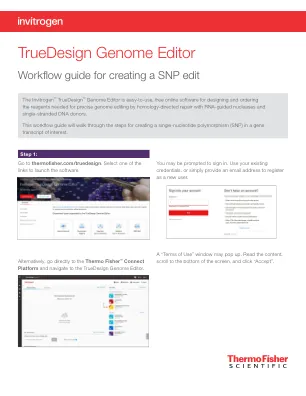
Truedesign基因组编辑器
要查看TALEN目标,请单击表的“ Talen”选项卡,并将显示每个Talen对的类似信息。TAL效应子核酸酶(TALEN)对。设计结果表中的绿色选中标记将指示推荐的技术。在Thermofisher.com/tal上了解有关Talen Technology的更多信息。