XiaoMi-AI文件搜索系统
World File Search SystemHSCS

可再生氢供应链
在全球范围内,能源系统正在经历向更可持续系统的过渡。根据氢路线图欧洲(FCH EU,2019年),由于其支持可持续性目标的能力,氢将在未来的能源系统中发挥重要作用,并将占未来总能量混合的13%。因此,正确的氢供应链(HSC)计划对于实现可持续过渡至关重要,特别是当使用可再生能源(可再生氢)的电力电解产生氢时。但是,由于可再生性HSC的运营特征,其计划很复杂。可再生氢供应可以多种多样:可以使用诸如风能和太阳能等续签的重点产生氢气,或者通过使用大容量的水力发电厂产生的电力来中心。同样,对氢的需求也可能是多种多样的,包括许多新的应用,例如用于燃料电池电动汽车和发电的燃料,工业过程中的原料以及用于建造的燃料。HSC由不同形式的各个阶段(生产,存储,分布和应用)组成,具有强大的相互依赖性,这进一步提高了HSC的复杂性。最后,HSC的规划取决于采用氢和市场发展的状态,以及技术的成熟程度,这两个因素都以高度不确定性为特征。直接适应HSC供应链(SC)计划的传统方法不足。我们仅关注可再生氢,因为它与未来的低碳经济相关。因此,在这项研究中,我们开发了一个具有相关计划任务的计划矩阵,利用系统的文献综述来应对HSC的特征。此外,从供应链管理的角度来看,我们概述了未来研究的议程,以支持可再生的HSC Devel Opment,考虑到可再生可再生性HSC采用和市场发展的不同阶段。

可再生氢供应链
全球范围内,能源系统正在向更可持续的系统过渡。根据《欧洲氢能路线图》(FCH EU,2019),氢能将在未来能源系统中发挥重要作用,因为它能够支持可持续发展目标,并将在未来占总能源结构的约 13%。因此,正确的氢供应链 (HSC) 规划对于实现可持续转型至关重要,尤其是当使用可再生能源(可再生氢)的电力通过水电解生产氢时。然而,由于可再生 HSC 的运行特性,其规划很复杂。可再生氢供应可以是多种多样的:氢可以利用风能和太阳能等可再生能源分散生产,也可以利用大量水力发电厂的电力集中生产。同样,对氢的需求也可以是多种多样的,有许多新的应用,例如燃料电池电动汽车和发电的燃料、工业过程中的原料以及建筑物供暖。 HSC 包括不同形式的各种阶段(生产、储存、分配和应用),相互依赖性很强,这进一步增加了 HSC 的复杂性。最后,HSC 的规划取决于氢气采用和市场开发的状况,以及技术的成熟度,这两个因素都具有高度不确定性。直接采用传统的 HSC 供应链 (SC) 规划方法是不够的。因此,在本研究中,我们开发了一个规划矩阵和相关规划任务,利用系统的文献综述来应对 HSC 的特点。我们只关注可再生氢,因为它与未来的低碳经济息息相关。此外,我们从供应链管理的角度概述了未来研究的议程,以支持可再生 HSC 的发展,同时考虑到可再生 HSC 的采用和市场开发的不同阶段。

安全高效的体内造血干细胞...
我们在恒河猴中测试了一种新的体内造血干细胞 (HSC) 转导/选择方法,使用 HSC 嗜性、整合性、辅助依赖性腺病毒载体 (HDAd5/35++),该载体旨在在红细胞 (RBC) 中表达人类 g -珠蛋白以治疗血红蛋白病。我们发现,HDAd5/35++ 载体在静脉注射到粒细胞集落刺激因子 (G-CSF)/AMD3100 动员的动物体内后优先转导 HSC,并且转导的细胞返回骨髓和脾脏。该方法耐受性良好,并且通常与静脉腺病毒载体注射相关的促炎性细胞因子的激活通过用地塞米松联合白细胞介素 (IL)-1 和 IL-6 受体阻滞剂进行预处理而成功减弱。使用我们基于 MGMT P140K 的体内选择方法,g-珠蛋白 +
创新方法提供基因编辑器ABE来治疗纳米技术与DNA折纸结合创新方法提供基因编辑器ABE来治疗纳米技术与DNA折纸结合
治疗方法很好,但是治疗的输送方法在临床上并不可行。接受HSC治疗的受体小鼠必须接受致命剂量的γ辐射,以消除大量的骨髓(其中包含绝大多数造血干细胞,HSC),从而带来了重大的安全问题。可能更希望寻求方法,而无需破坏骨髓中HSC的原始菌落。一种方法是制定腺嘌呤碱基编辑器(ADE),并通过静脉内途径将其传递到骨髓中,以实现体内编辑。图1显示了有关治疗小鼠的一些统计数据,包括β球蛋白的百分比,细胞形态等。可以清楚地看出,经过治疗的小鼠的细胞已恢复正常的形态,证明了这种治疗的有效性。

造血干细胞识别射线后
辐射暴露尤其损害造血系统的细胞,诱导全血管减少症和骨髓衰竭。对这些过程的研究,以及开发治疗以防止造血损伤或增强辐射暴露后的恢复,通常需要在辐射后早期对骨髓细胞进行分析。虽然流式细胞术方法的表征很好地鉴定和分析了非辐照环境中的骨髓种群,但在处理辐照组织时会出现多种并发症。是辐射引起的C-KIT丧失,这是小鼠原始造血种群传统门控的中心标记。这些包括造血干细胞(HSC),这些干细胞是血液重建和终身骨髓功能的核心,并且是这些研究中分析的重要靶标。本章概述了HSC识别和分析的技术。
利用 NxT 的最佳技术进行转染 - Rhenium Bio
图 3. Neon NxT 重悬基因组编辑缓冲液在不同细胞类型和靶标的 CRISPR-Cas9 基因组编辑实验中的表现。靶标位点包括 Jurkat 和 K562 细胞的 ACTN、活化原代 T 细胞的 TRAC、HSC 的 B2M 和原代 NK 细胞的 AAVS1。细胞在 10 µL 或 100 µL 反应中进行电穿孔。(A) GFP 供体 DNA 敲入效率报告为 GFP 阳性细胞的百分比。(B) GFP 供体 DNA 敲入后的细胞活力。(C) 敲除效率报告为与未处理对照相比特定靶标位点的减少百分比。对于原代 NK 细胞,通过基因组切割检测 (GCD) 测定确定的插入/缺失效率 (%) 可作为敲除效率的指标。(D) 敲除细胞的电穿孔后活力。

标题 经过基因编辑的 CD34+ 细胞源自人类 iPS 细胞(在体内但不是体外),可植入并分化为抗 HIV 细胞
iPS 细胞 | CCR5 | HIV 抗性 | 基因编辑 | 畸胎瘤 近期 HIV 研究的主要目标是开发一种“治愈”这种病毒感染的方法,避免终身接受抗逆转录病毒疗法 (ART)。实现这一目标的方法之一是删除或突变编码促进 HIV 感染和传播的蛋白质的基因。这一策略的一个有吸引力的候选基因是 Ccr5 基因,该基因突变导致 32 bp 缺失,已被证明与天然保护免受 HIV 感染和疾病有关 (1, 2)。Ccr5 基因编码 CCR5,这是一种人类细胞表面趋化因子受体,是 HIV 附着和感染细胞的辅助受体 (3, 4)。Ccr5 等位基因的 32 bp 缺失导致 CCR5 受体的截短异构体 CCR5 Δ 32,它不在细胞表面表达。因此,病毒进入细胞被阻止 (5)。诱导性多能干 (iPS) 细胞 (6) 能够分化为 CD34 + 造血干细胞 (HSC) (7),因此可以重建完整的免疫系统 (8, 9)。因此,这些 iPS 细胞是基因工程的首选目标。我们小组和其他小组已经证明,由健康个体 (10) 和接受 ART 治疗的 HIV 感染患者 (11) 的外周血单核细胞 (PBMC) 产生的 iPS 细胞可以经过基因编辑,使其 Ccr5 基因的野生型等位基因携带 Ccr5 Δ 32 突变 (12, 13)。值得注意的是,使用 CRISPR/Cas9 技术,可以修改 Ccr5 基因,使其具有与对 R5 嗜性病毒的抵抗力相关的天然 Δ 32 变体等位基因。此外,虽然截短的 CCR5 Δ 32 蛋白不存在于细胞表面,但它仍然表达,因此可能具有其他重要的生理作用(14-17)。我们已经证实,基因改造的 Ccr5 Δ 32 iPS 细胞可以在体外分化为 CD34 + HSC(10,18)。在适当的细胞培养条件下,它们可以产生各种
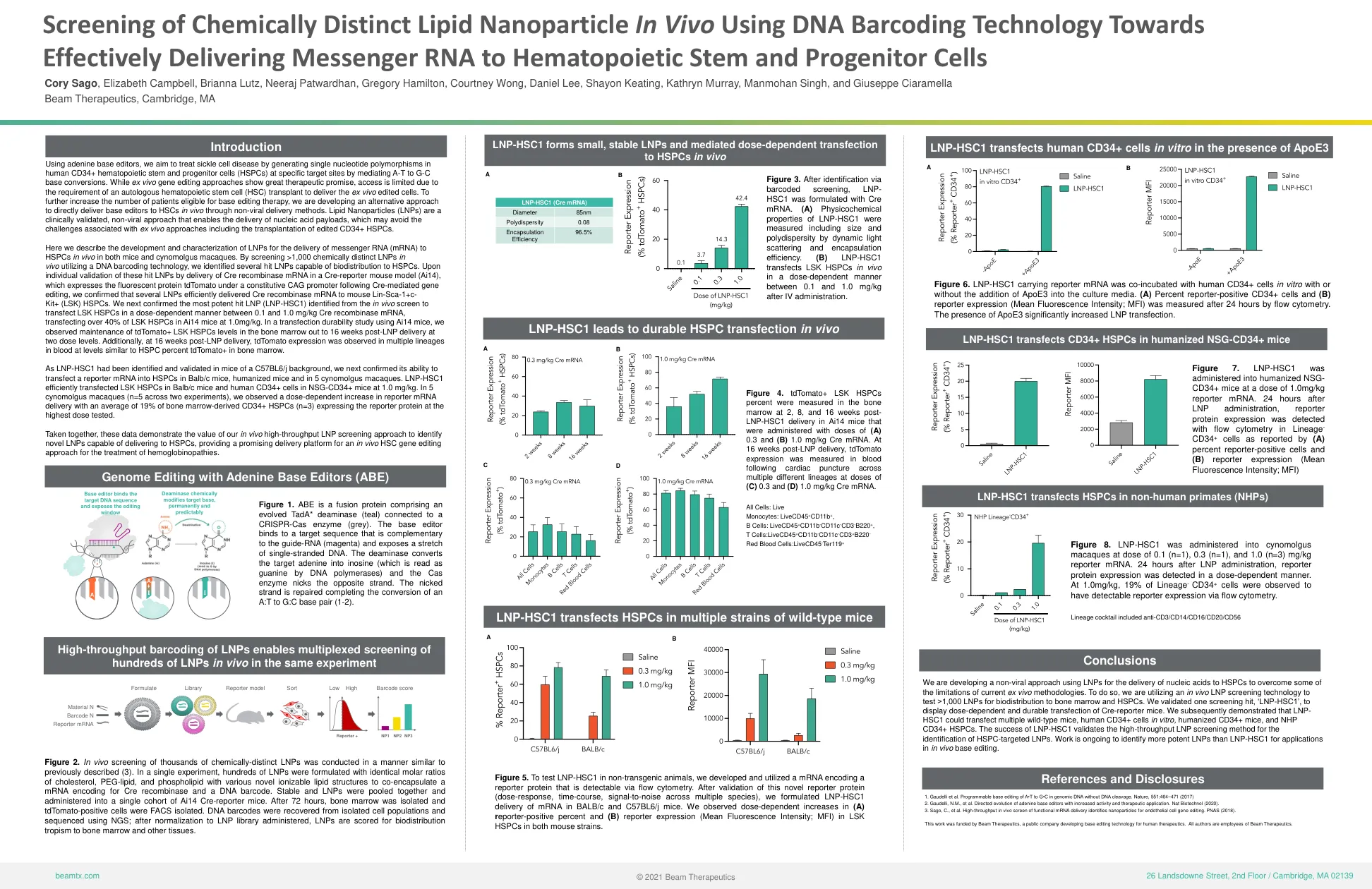
202111-ash-screening-of-chemically-distinct-lnp-in-vivo。......
我们的目标是利用腺嘌呤碱基编辑器,通过介导 AT 到 GC 碱基的转化,在特定靶位点的人类 CD34+ 造血干细胞和祖细胞 (HSPC) 中产生单核苷酸多态性,从而治疗镰状细胞病。虽然离体基因编辑方法显示出巨大的治疗前景,但由于需要自体造血干细胞 (HSC) 移植来递送离体编辑的细胞,因此获取途径有限。为了进一步增加有资格接受碱基编辑治疗的患者数量,我们正在开发一种替代方法,通过非病毒递送方法将碱基编辑器直接递送到体内的 HSC。脂质纳米颗粒 (LNP) 是一种经过临床验证的非病毒方法,可以递送核酸有效载荷,从而可以避免与离体方法相关的挑战,包括移植编辑的 CD34+ HSPC。


从造血干细胞到血小板
摘要:血小板是主要在骨髓中产生的巨核细胞的末端后代,在血液稳态,凝结和伤口愈合中起关键作用。传统上,巨核细胞和血小板被认为是由多个离散的祖细胞(HSC)引起的,这些造血细胞(HSC)通过多个离散的祖细胞,并具有连续的,谱系限制的差异步骤。然而,最近的研究挑战了这种观点,该研究表明(1)某些HSC克隆有偏见和/或仅限于血小板谱系,(2)并非所有血小板都会产生遵循“典型”巨核细胞分化路径的造血性巨核细胞,以及(3)血小板输出量是稳定稳定性稳定稳定性稳定型Hematopoiesisis septecteale septectea。在这里,我们特别研究了体内谱系追踪研究提供了血小板生成的途径的证据,并研究了各种中间祖细胞群体的参与。我们进一步确定了确定这些可能替代途径的存在,角色和动力学所需的挑战。