XiaoMi-AI文件搜索系统
World File Search SystemKD

kurukshetra大学kurukshetra
1。Arthur Beiser的现代物理学概念,McGraw Hill Education。2。现代物理(第2版),S.L。Kakani和Shubhra Kakani,Viva Books,新德里。 3。 半导体设备 - S.M .Sze的物理和技术,Wiley(1985)4。 激光和非线性光学元件B.B.Laud。 ,Wiley Eastern Limited(1985)5。 A.K. Sharma的半导体电子产品,新时代国际出版商(1996)6。 肯尼斯·克兰 辐射检测和测量:G.F. Knoll(纽约Wiley)(2000)8。 Verma和Srivastava:固态物理学的晶体学9. rajnikant;固态物理学,威利印度,2011年。 10。 J.C. Anderson,KD。 Leaver,R.D。 Rawlings和J.M. 亚历山大,材料科学,第四Kakani和Shubhra Kakani,Viva Books,新德里。3。半导体设备 - S.M .Sze的物理和技术,Wiley(1985)4。激光和非线性光学元件B.B.Laud。,Wiley Eastern Limited(1985)5。A.K. Sharma的半导体电子产品,新时代国际出版商(1996)6。肯尼斯·克兰辐射检测和测量:G.F. Knoll(纽约Wiley)(2000)8。Verma和Srivastava:固态物理学的晶体学9.rajnikant;固态物理学,威利印度,2011年。10。J.C. Anderson,KD。 Leaver,R.D。 Rawlings和J.M. 亚历山大,材料科学,第四J.C. Anderson,KD。Leaver,R.D。Rawlings和J.M.亚历山大,材料科学,第四

Andrew C. Kruse Research Isperials教育学术...Andrew C. Kruse Research Isperials教育学术...
Rajashankar K, Anderson KS, HR Baxter, Blacklow SC, TJ Boggon, Bonvin AM, Borek D, Brett TJ, Chaflisch A, Chazin WJ, KD Corbet, Fan, Fan QR, Ferré-D'Amare AR, Christopher From J, Garcia KC, Golden R, Gong P, Harrison SC, Heldwein EE, Jia Z, the RJ Cruise,Cross AC,Kvansakul M,McLellan JS,Modidi和Nam,Otwinski Z,Pai EF,Pereira PJ,CS Raman CS,Rapport CS,Rapport CS,报告。 TA,Roll-Mecak A,Rosen MK,Redrone G,Schwartz TU,Shamoo Y,Sandermann H,Tao YJ,NH Tolia,Tsodikov OV,Westover KD,Wu H,P。数据出版(2016)NAT。公社。7,10882

CK SUMESH 博士
4. Pratik Pataniya、Chetan K. Zankat、MohitTannarana、CK Sumesh、Som Narayan、GK Solanki、KD Patel、VM Pathak、Prafulla K. Jha “由 WSe2 纳米点功能化的纸基柔性光电探测器” ACS Appl. Nano Mater.2,5, 2758-2766 (2019)。5. Abhishek Patel、Pratik Pataniya、GK Solanki、CK Sumesh、KD Patel、VM Pathak “n-VO 2 /n-MoSe 2 异质结二极管的制造、光响应和温度依赖性” Superlattices and Microstructures 130, 160-167 (2019)。 6. CK Sumesh “MX 2(M = Mo,W;X = S,Se)/Si异质结器件的温度相关电子电荷传输特性”材料科学杂志:电子材料;30,4117–4127(2019 年)。7. CK Sumesh “纳米结构太阳能电池中的高效光子管理:2D 层状过渡金属二硫属化物半导体的作用”太阳能材料和太阳能电池 192 16–23(2019 年)。8. CK Sumesh 和 Kinnari Parekh “纳米催化物理化学吸附和有机染料降解”Pramana – 物理学杂志(2019 年)92:87 DOI:10.1007/s12043-019-1760-0(2019 年)。 9. SanniKapatel、CK Sumesh,“两步简便制备 MoS2·ZnO 纳米复合材料作为亚甲蓝(染料)降解的有效光催化剂”15,119–132 (2019)。10. Pratik Pataniya、GK Solanki、Chetan K. Zankat、MohitTannarana、CK Sumesh、KD Patel、VM Pathak,“n-

自噬激活可以部分挽救蛋白酶体功能障碍介导的心脏毒性
图 1 心脏靶向(Gal4 Τ inC Δ 4 )蛋白酶体 Pros β 5 基因的 KD 导致蛋白质组不稳定和线粒体数量减少。 (a) Pros β 5 siRNA 后心脏组织中 Pros β 5 基因的相对表达(与对照相比)。 (b, c) Pros β 5 RNAi(与对照相比)果蝇心脏组织中相对 (%) 26S 蛋白酶体活性 (b) 和 ROS 水平 (c)。 (d) Pros β 5 KD 后果蝇心脏组织中蛋白质组泛素化 (Ub) 和羰基化 (DNP) 的免疫印迹分析。 (e) CLSM 观察用 LysoTracker 染色的 Pros β 5 RNAi(与对照相比)果蝇心管(e1)、LysoTracker 定量(e2)和使用溶酶体标记物抗 Lamp1(e3)进行免疫印迹分析。(f) 所示基因型果蝇心脏组织中蛋白酶活性的相对(%)。(g) blw/ATP5A 免疫荧光染色后,CLSM 可视化所示果蝇品系心脏组织中的线粒体;细胞核用 DAPI 复染。(h) Pros β 5 KD 后,所示基因型分离心脏组织中所示线粒体基因的相对表达水平(与对照相比)。在 (a, h) 中,基因表达与相应对照作图;使用 RpL32/rp49 基因作为 RNA 输入参考。 (d)和(e3)中的 Gapdh 和 Actin 探测分别用作蛋白质输入参考。p 值采用非配对 t 检验计算。条形图,± SD(n ≥ 3);* p < 0.05;** p < 0.01
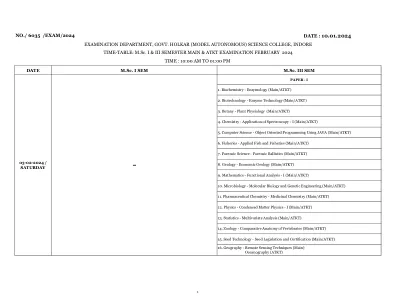
时间表:硕士I&III学期主和ATKT考试2024年2月
UKS&1- TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK TK ICB〜;ØeGSRQ{K@LACAF/KR f'kd laidz


PHF19 抑制作为多发性骨髓瘤的治疗靶点
表观遗传失调越来越多地被认为是多发性骨髓瘤 (MM) 的一个致病因素。特别是 H3 赖氨酸 27 (H3K27me3) 的三甲基化,它由多梳抑制复合物 2 (PRC2) 的亚基 PHD 指蛋白 19 (PHF19) 催化,最近已被证明是 MM 致瘤性的关键介质。在 MM 中 PHF19 的过度表达与更差的临床结果有关。然而,虽然有越来越多的证据表明 PHF19 过度表达在 MM 致癌作用中起着关键作用,但下游机制仍有待阐明。在当前的研究中,我们使用 PHF19 的功能性敲低 (KD) 来研究 PHF19 的生物学作用,并表明 PHF19KD 会导致体外和体内肿瘤生长减少。 PHF19 KD 后,bcl2、myc 和 EGR 等主要癌症因子的表达降低,进一步强调了 PHF19 在 MM 生物学中的作用。此外,我们的研究结果还强调了 PHF19 过表达对预后的影响,这与生存率下降显著相关。总体而言,我们的研究强调了这样一个前提:针对 PHF19-PRC2 复合物将为新型 MM 疗法开辟道路。
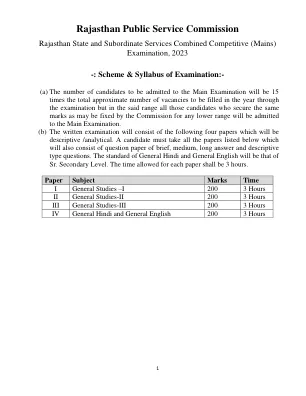
拉贾斯坦邦公共服务委员会
Hkkx v& ( vad 50 ) • `Fkd~ djuk • izR;; & fn,gq,izR;;ksa ls'kCn cukuk vkSj'kCnksa esa ls ewy'kCn,oa izR;; i`Fkd~ djuk • i;kZ;okph 'kCn • foykse 'kCn • leJqr fHkUukFkZd 'kCn&fn, gq, 'kCn&;qXe dk vFkZ&Hksn • okD;ka'k ds fy, lkFkZd 'kCn • 'kCn •qn •qD~k; 'kqn~f/k • eqgkojs& eqgkojxa dk okD; esa iz;ksx ls vFkZ Li"V • dgkor@yksdksfDr&okD; esa iz;ksx ls vFkZ Li"V • ikfjHkkf"kd 'kCnkoyh& iz'kklu ls lacaf/kr vaxzsth 'kCnksa ds lekukFkZ ik fjUnhkkd 'Hkkf"

核酸研究
摘要MHC I类鼠和β-2-微球蛋白基因在胚胎癌(EC)细胞中是沉默的,但在分化这些细胞时会诱导。我们先前表明,位于H-2KB基因启动子中的增强子样序列在F9和PCC3细胞中是非功能的。我们先前已经从小鼠T细胞系中纯化了48 kD蛋白(KBFI),该细胞系与该增强子中的腔序列序列结合,并与Beta-2-微球蛋白基因启动子中的类似序列结合。我们在这里解散第二蛋白(Kbf2,58 kd)的纯化,该蛋白也与该序列结合。虽然两种活性都存在于分化细胞中,但在未分化的EC细胞中不存在KBF1结合活性,在未分化的EC细胞中,腔液序列没有增强剂活性。分化后,诱导KBF1结合活性,而palindromic序列作为增强子变得活跃。因此,在未分化的EC细胞中缺乏KBF1活性至少部分原因是缺乏在这些细胞中H-2 I类和β-2-微球蛋白基因表达的表达,并表明KBF1活性在分化过程中受到调节。
