XiaoMi-AI文件搜索系统
World File Search SystemLineage

了解细胞谱系,细胞多样性和人脑通过干细胞模型的演变
大脑皮层由谷氨酸能和GABA能神经元亚型多样化,这些亚型显示出特定的电生理,分子,形态学和Hodologicy特性。兴奋性投射神经元来自发育中的皮质内的背祖细胞,而GABA能抑制性抑制性中间神经元是在尾和中间神经节eminence(CGE和MGE分别)以及前尾部和中间的神经节感染性中产生的,以及在preoptic区域(POA)。皮质神经元被组织成层,这些层是由跨时间的内而外的迁移模式产生的。早期出生的神经元填充了深层,而后长的神经元侵入皮质的上层。给定兴奋性皮质神经元的层状,分子和实际身份定义了其在大脑其余部分内的特定连通性模式[1]。单细胞转录组研究强调了在人类和小鼠中部分保守的皮质兴奋性神经元的显着多样性[2]。在哺乳动物之间也保守了不同神经亚型的顺序产生。然而,根据物种特异性类型的神经祖细胞的存在以及祖细胞池扩张,神经发生和最终神经元成熟的特定时机可能会有所不同。另外,神经元亚型的层分布[2]和不同大脑区域中每一层的相对大小在灵长类动物和啮齿动物中都有变化[3]。

单个细胞中的关节谱系,转录组和表观基因组分析的高克隆条形码多样性的小鼠模型
细胞谱系历史及其分子状态编码组织发育和稳态的基本原理。当前的谱系录制小鼠模型的条形码多样性有限,单细胞谱系覆盖范围较差,从而排除了它们在由数百万个细胞组成的组织中的使用。在这里,我们开发了Darlin,这是一种改进的CAS9条形码小鼠系,它利用末端脱氧核苷酸转移酶(TDT)来增强30个CRISPR目标位点的插入事件,稳定地整合到3个不同的基因组基因座中。darlin是可诱导的,估计有〜10 18个层次条形码,并可以检测约60%的剖面单细胞中可用的条形码。使用Darlin,我们检查了发育中的造血干细胞(HSC)中的命运启动,并揭示了HSC迁移的独特特征。此外,我们为单个细胞中的共同介绍了一种方法来共同介绍DNA甲基化,染色质可及性,基因表达和谱系信息。darlin将在各种组织和生理环境中对谱系关系及其分子特征进行广泛的高分辨率研究。

一种简单的方法,可以在原位杂交后识别神经板阶段的海囊脑谱系细胞
克莱尔·哈德森(Clare Hudson)。一种简单的方法,可以在原位杂交后在神经板阶段识别海腹脑谱系细胞。Simon G. Sprecher。大脑发育。方法和协议,施普林格,第325-345页,2020年,《分子生物学中的方法》,978-1-4939-9731-2。10.1007/978-1-4939-9732-9_18。hal-02322828

泰国 SARS-CoV-2 基因组监测显示,存在混合输入种群、本地谱系扩张和 ORF7a 截短的病毒
。CC-BY-NC-ND 4.0 国际许可 它是根据作者/资助者提供的,他已授予 medRxiv 永久展示预印本的许可。(未经同行评审认证)
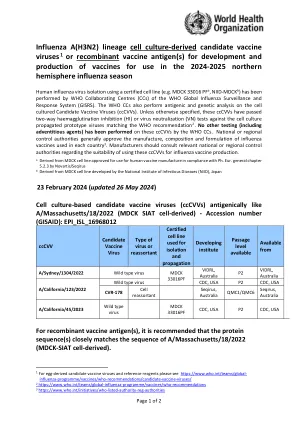
流感A(H3N2)谱系细胞培养衍生的候选疫苗病毒1或重组疫苗抗原,用于疫苗的开发和生产
流感 A(H3N2) 谱系细胞培养衍生的候选疫苗病毒 1 或重组疫苗抗原,用于开发和生产用于 2024-2025 年北半球流感季节的疫苗。世卫组织全球流感监测和应对系统 (GISRS) 的世卫组织合作中心 (CC) 已使用认证细胞系(如 MDCK 33016 PF a、NIID-MDCK b)分离人流感病毒。世卫组织 CC 还对细胞培养的候选疫苗病毒 (ccCVV) 进行抗原和基因分析。除非另有说明,这些 ccCVV 已通过针对符合世卫组织建议 2 的细胞培养繁殖原型病毒的双向血凝抑制 (HI) 或病毒中和 (VN) 测试。世卫组织 CC 尚未对这些 ccCVV 进行任何其他测试(包括外来因子)。国家或地区监管部门通常会批准每个国家使用的流感疫苗的生产、成分和配方 3 。制造商应咨询相关国家或地区监管部门,了解使用这些 ccCVV 进行流感疫苗生产的适用性。

不完整的谱系分类解释了在四种西欧蚱hoppers(Orthoptera:acrididae:chorthippus)的辐射中DNA条形码低的性能
Benoit Nabholz。不完整的谱系分类解释了四种西欧蚱hoppers的辐射中DNA条形码的低性能(Orthoptera:Acrididae:Chorthippus)。林尼社会生物学杂志,2023年,10.1093/biolinnean/blad106。hal-04192161
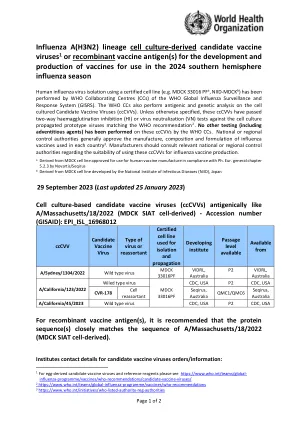
流感A(H3N2)谱系细胞培养衍生的候选疫苗病毒1或重组疫苗抗原,用于疫苗的开发和生产
流感 A(H3N2) 谱系细胞培养衍生的候选疫苗病毒 1 或重组疫苗抗原,用于开发和生产用于 2024 年南半球流感季节的疫苗。世卫组织全球流感监测和应对系统 (GISRS) 的世卫组织合作中心 (CC) 已使用认证细胞系(如 MDCK 33016 PF a、NIID-MDCK b)分离人流感病毒。世卫组织 CC 还对细胞培养的候选疫苗病毒 (ccCVV) 进行抗原和基因分析。除非另有说明,这些 ccCVV 已通过针对符合世卫组织建议 2 的细胞培养繁殖原型病毒的双向血凝抑制 (HI) 或病毒中和 (VN) 测试。世卫组织 CC 尚未对这些 ccCVV 进行任何其他测试(包括外来因子)。国家或地区监管部门通常会批准每个国家使用的流感疫苗的生产、成分和配方 3 。制造商应咨询相关国家或地区监管部门,了解使用这些 ccCVV 进行流感疫苗生产的适用性。

单核RNA-Seq表征了成人人类亚依赖型区域中神经元谱系的细胞类型,并揭示了年龄
亚依赖型区(SEZ),也称为室室区(SVZ),构成了在产后生活期间持续存在的神经源性生殖位。在人类中,生命第一年后,经济特区的神经源性潜力下降。然而,发现茎和祖细胞标记的研究突出了成年人类经济特区祖细胞的神经源能力,神经源性活性增加在病理条件下发生。在本研究中,成年人类经济特区的完整细胞壁细分的特征是单核RNA测序,并比较了四名青年(16-22岁)和四个中年成年人(44-53岁)。我们确定了11个细胞簇,包括表达神经干细胞(NSC),神经细胞,未成熟神经元和少突胶质细胞祖细胞细胞的标记基因的簇。两个年龄组之间NSC和神经细胞簇的相对丰度没有差异,这表明SEZ NSC的库在该年龄范围内不会下降。中年少突胶质细胞祖细胞和小胶质细胞的相对丰度降低,表明人类经济委的细胞组成在青年和成年之间被重塑。与中年相比,在包括NSC在内的不同细胞类型(包括NSC)中,与神经系统发育相关的基因的表达较高。这些转录的变化表明,在中年龄段下降的年轻人中,经济特区中的中枢神经系统持续的可塑性。
磷酸化蛋白质组学分析揭示了人类骨髓衍生的基质干细胞的成骨细胞谱系的定义遗传程序
骨髓 - 衍生的间充质干细胞(MSC)在其小众中存在的信号刺激后分化为成骨细胞。由于与MSC的成骨细胞(OB)分化相关的全局信号传导级联反应没有很好地定义,因此我们使用定量质谱法来描述人类MSC蛋白质组和磷酸化型的变化。6252蛋白和15,059个磷光位点的时间曲线表明至少两个不同的信号传导波:刺激后30至60分钟内的一个峰值在30至60分钟内峰值,在24小时后进行了第二次升高。除了在早期MSC分化过程中提供蛋白质组和磷酸蛋白质组动力学的全面视图外,我们的分析还确定了丝氨酸/苏氨酸蛋白激酶D1(PRKD1)在OBS中的关键作用。在OB分化开始时,PRKD1通过触发组蛋白脱乙酰基酶HDAC7的磷酸化和核排除来启动促稳态转录因子Runx2的激活。
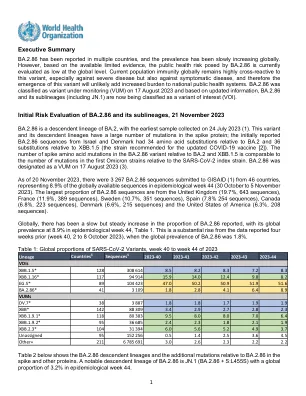
BA.2.86 及其亚谱系的初步风险评估,2023 年 11 月 21 日 BA.2.86 是 BA.2 的后代谱系,最早采集的样本
BA.2.86 及其亚谱系的初步风险评估,2023 年 11 月 21 日 BA.2.86 是 BA.2 的后代谱系,最早的样本采集于 2023 年 7 月 24 日 (1)。该变体及其后代谱系的刺突蛋白有大量突变;最初报告的来自以色列和丹麦的 BA.2.86 序列相对于 BA.2 有 34 个氨基酸替换,相对于 XBB.1.5(推荐用于更新的 COVID-19 疫苗的菌株 [2])有 36 个氨基酸替换。BA.2.86 变体相对于 BA.2 和 XBB.1.5 的刺突氨基酸突变数量与第一批 Omicron 菌株相对于 SARS-CoV-2 指标菌株的突变数量相当。BA.2.86 于 2023 年 8 月 17 日被指定为 VUM (3)。截至 2023 年 11 月 20 日,共有来自 46 个国家的 3 267 个 BA.2.86 序列提交给 GISAID (1),占流行病学第 44 周(2023 年 10 月 30 日至 11 月 5 日)全球可用序列的 8.9%。BA.2.86 序列中占比最大的国家是英国(19.7%,643 个序列)、法国(11.9%,389 个序列)、瑞典(10.7%,351 个序列)、西班牙(7.8%,254 个序列)、加拿大(6.8%,223 个序列)、丹麦(6.6%,215 个序列)和美国(6.3%,208 个序列)。在全球范围内,已报告的 BA.2.86 比例缓慢但稳定地增加,流行病学第 44 周的全球流行率为 8.9%,见表 1。与四周前(第 40 周,2023 年 10 月 2 日至 8 日)报告的数据相比,这是一个大幅增长,当时 BA.2.86 的全球流行率为 1.8%。表 1:2023 年第 40 周至第 44 周 SARS-CoV-2 变体的全球比例