XiaoMi-AI文件搜索系统
World File Search SystemNGG

Edit-R™ CRISPR 设计工具
图 1. Edit-R™ CRISPR 设计工具如何选择针对人类基因 PPIB 的 20 个核苷酸序列的示例。目标序列可以位于基因组 DNA 的任一链上,只要它处于 5' 到 3' 方向,并且该链的 3' 端有一个 NGG PAM(使用默认的 S. pyogenes Cas9 时)。Cas9 核酸酶将在 NGG PAM 上游三个核苷酸的位置切割 DNA 的两条链。建议选择完全位于编码基因早期组成外显子内的靶位点,但 Edit-R™ CRISPR 设计工具将返回整个编码区域的结果,因此如果需要,可以靶向特定的外显子或蛋白质结构域。
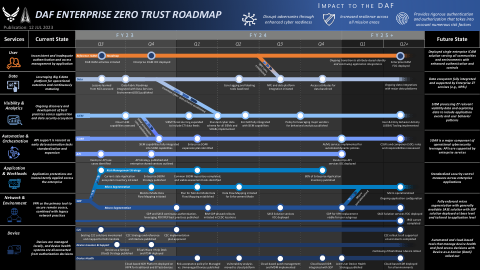
DAF 企业零信任路线图
删除了“PACAF SDP 试点已部署”里程碑,以反映这些试点停止的最新决定;在 2023 财年第四季度末和 2025 财年第二季度及以后分别添加了额外的里程碑(来自原始政策泳道),以显示 SASE 解决方案 FOC 的部署时间表;在“BIM SDP 分阶段推出已启动”里程碑中添加了澄清语言,以澄清 NGG 执行的初始阶段将从 CEDC 位置开始,然后再过渡到特殊用途处理节点 (SPPN)

利用-QM-MM和分子型模拟 -
图2。DNA,SGRNA和蛋白质相互作用(a)(a)匹配的SPCAS9和(b)MM5-SPCAS9聚焦HNH催化位点和PAM(NGG)区域。(C&D)显示了匹配的和MM5的不同视图,从而缩放了PAM远端和RUVC区域相互作用。T-DNA,NT-DNA和SGRNA分别为颜色的洋红色,黄色和浅蓝色。SPCAS9,HNH和RUVC的两个核酸酶结构域以白色和深蓝色显示。

CRISPR-Cas9 载体
CRISPR/Cas9系统已被广泛应用于基因组编辑,包括基因破坏、定点诱变、表观遗传调控等。化脓性链球菌(SpCas9)是目前最常用的Cas9蛋白。通过SpCas9进行基因组编辑需要在靶位点有一个“NGG”原型间隔区相邻基序(PAM)序列,这限制了CRISPR/Cas9系统的编辑范围。为了扩大编辑位点的范围和优化编辑特异性,各种SpCas9突变体已被研究并成功应用于CRISPR系统。

通过PCR协议形式的基因分型
等位基因描述:使用优化的CRISPR CAS9 KI技术创建了鼠标R565Q模型,该技术利用核糖核蛋白(RNP)在存在带有所需KI的合成单链DNA修复模板的情况下创建了。zygotes被电穿孔,并通过同源性修复(HDR)筛选后后代,以确定正确靶向等位基因的存在。Ki核苷酸破坏了PAM位点(NGG),为4和5 bp,与裂解位点并设计为SSODN,以保护HDR衍生的基因座免受CAS9自动切割。关键后代。

利用 TadA 的下一代胞嘧啶碱基编辑器
• 脱氨酶的定向进化 • PAM 变体碱基编辑器 • 定向进化 Cas9 以创建用于 BE 的非 NGG PAM 变体 • 密码子、NLS 和接头优化 • 环状置换体和镶嵌碱基编辑器 • DNA 脱靶评估 • RNA 脱靶评估 • 旁观者编辑最小化 • 引导 RNA 工程 • 离体和体内 BE 递送 • 最小化脱靶活性的工程 BE • HSC、肝细胞和 T 细胞的离体碱基编辑 • ABE 的低温电子显微镜结构 • 小鼠体内碱基编辑 • 非人类灵长类动物体内编辑

Issatchenkia Orientalis
图1。SD108中全基因组整合位点的硅筛选算法算法。 (a)用于ICAS9介导的整合的基因基因座中的GRNA。 扫描基因组以获取“ NGG” PAM以获得指南RNA库。 筛选GRNA以最大程度地减少潜在的脱靶,并根据其基因组位置过滤。 (b)纳入各种因素以优先考虑基因组基因局进行实验筛查。 GRNA及其相应的同源臂是根据寡核苷酸合成和质粒克隆标准来完善的。 设计规则是通过避免调节元素的破坏和包括基因本质信息的中断来确保应变稳定性的,而基因密度则是添加基因密度作为开放染色质的代理。 转录组数据纳入了接近转录活性基因的选择位置。算法。(a)用于ICAS9介导的整合的基因基因座中的GRNA。扫描基因组以获取“ NGG” PAM以获得指南RNA库。筛选GRNA以最大程度地减少潜在的脱靶,并根据其基因组位置过滤。(b)纳入各种因素以优先考虑基因组基因局进行实验筛查。GRNA及其相应的同源臂是根据寡核苷酸合成和质粒克隆标准来完善的。设计规则是通过避免调节元素的破坏和包括基因本质信息的中断来确保应变稳定性的,而基因密度则是添加基因密度作为开放染色质的代理。转录组数据纳入了接近转录活性基因的选择位置。

1 用于多拷贝基因整合的着陆垫系统...
图 1. SD108 中全基因组整合位点的计算机筛选算法。(A)选择基因间位点中的 gRNA 进行 iCas9 介导的整合。扫描基因组中的“NGG”PAM 以获得向导 RNA 文库。筛选 gRNA 以尽量减少潜在的脱靶,并根据其基因组位置进行过滤。(B)结合各种因素对实验筛选的基因组位点进行优先排序。根据寡核苷酸合成和质粒克隆标准对 gRNA 及其相应的同源臂进行改进。实施设计规则以确保菌株稳定性,避免破坏调控元件并包括基因必需性信息,同时添加基因密度作为开放染色质的代理。结合转录组学数据来选择靠近转录活性基因的位点。

通过腺嘌呤碱基编辑遗传性视网膜疾病的成年小鼠的视觉功能恢复
胞嘧啶碱基编辑器和腺嘌呤碱基编辑器(ABE)可以可预测地校正点突变,并且独立于CAS9诱导的双链DNA断裂(这会导致实质性的indel形成)和同源性指导的修复(通常会导致较低的编辑效率)。在此,我们在成年小鼠中表明,在RPE65基因中,态慢性病毒的下视网膜下注射表达ABE和单一指导RNA,靶向从RPE65基因进行的无义突变纠正了致病性突变,可纠正效率高达29%的效率,并在indel和oft oft oftarget的突变中均具有最小的效率,但均具有29%的效率,并且是不可或缺的效率。主题。ABE处理的小鼠显示了恢复的RPE65表达和类视黄素异构酶活性,以及视网膜和视觉功能的接近正常水平。我们的发现激发了对

通过腺嘌呤碱基编辑恢复患有遗传性视网膜疾病的成年小鼠的视觉功能
胞嘧啶碱基编辑器和腺嘌呤碱基编辑器 (ABE) 可以可预测地纠正点突变,并且不受 Cas9 诱导的双链 DNA 断裂(导致大量插入/缺失形成)和同源定向修复(通常导致低编辑效率)的影响。本文,我们在成年小鼠中表明,视网膜下注射表达 ABE 的慢病毒和针对 Rpe65 基因中新生无义突变的单向导 RNA 可以纠正致病突变,效率高达 29%,并且插入/缺失和脱靶突变的形成最少,尽管没有典型的 NGG 序列作为原间隔区相邻基序。经 ABE 处理的小鼠显示恢复的 RPE65 表达和类视黄酸异构酶活性,以及接近正常水平的视网膜和视觉功能。我们的发现促使进一步测试 ABE 以用于