XiaoMi-AI文件搜索系统
World File Search SystemPKC

PI3K/mTOR 是一种可治疗的基因...
弥漫性中线胶质瘤 (DMG),包括脑干中诊断出的肿瘤(弥漫性内在性脑桥胶质瘤;DIPG),都是缺乏有效治疗的致命性脑肿瘤。CRISPR/Cas9 功能丧失基因缺失筛选分析确定 PIK3CA 和 MTOR 是患者衍生的 DIPG 模型中可靶向的分子依赖性,凸显了血脑屏障渗透性 PI3K/Akt/mTOR 抑制剂 paxalisib 的治疗潜力。在人类等效最大耐受剂量下,用 paxalisib 治疗的小鼠经历了全身葡萄糖反馈和胰岛素水平升高,与使用 PI3K 抑制剂的患者相当。为了利用遗传依赖性并克服耐药性,同时保持依从性和治疗效果,我们将 paxalisib 与抗高血糖药物二甲双胍联合使用。二甲双胍恢复了葡萄糖稳态并降低了体内胰岛素受体的磷酸化,这是 PI3K 抑制剂耐药的常见机制,从而延长了原位模型的生存期。用 paxalisib 治疗的 DIPG 模型增加了钙激活的 PKC 信号传导。脑渗透性 PKC 抑制剂 enzastaurin 与 paxalisib 联合使用,协同延长了多种原位患者来源和免疫功能正常的同基因移植模型的生存期;与二甲双胍和标准护理放射治疗联合使用可增强益处。使用空间转录组学和 ATAC-Seq 评估治疗适应性,确定髓鞘形成和肿瘤免疫微环境串扰的变化。总的来说,这项研究确定了我们认为具有临床意义的 DIPG 治疗组合策略。

XL在小场上的理论分析
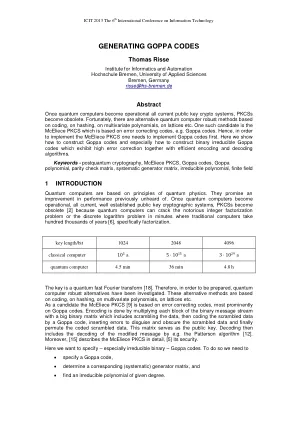
摘要。XL首先是为了解决有限范围内确定的或过度确定的方程式系统,作为对多个密码系统的“代数攻击”。通过此类攻击(包括流密码)对基本的密码分析一直存在稳定的公告(例如,toyocrypt),PKC和更具争议性的密码(AES/Rijndael和Serpent)。对XL的先前讨论通常在模拟中很重,这当然很有价值,但我们希望更多地关注理论,因为理论和模拟必须互相验证,并且有些细微差别不易与模拟。在最近的这个方向上付出了更多的努力,但其中大部分仅限于大小的大基地,通常等于

和心脏成纤维细胞的磷酸化 - 磷酸化 - la trobe
心肌细胞和成纤维细胞蛋白质组景观的抽象病理重编程驱动心脏纤维化的起始和进展。尽管功能障碍性心肌细胞的分泌成为病理成纤维细胞重编程的重要驱动力,但我们对下游分子娱乐体的理解仍然有限。在这里,我们表明心脏成纤维细胞激活(αSMA +)和由TGFβ刺激的心肌细胞的分泌介导的氧化应激与其蛋白质组和磷酸蛋白酶景观的深刻重新编码有关。在成纤维细胞全局蛋白质组中,蛋白质的失调引起的失调与细胞外基质,蛋白质定位/代谢,KEAP1-NFE2L2途径,溶酶体,碳水化合物,碳水化合物的代谢和转录调节。激酶底物富集分析磷酸肽在此重塑过程中激酶(CK2,CDK2,PKC,GSK3B)的潜在作用。 我们验证了酪蛋白激酶2(CK2)在分泌理论治疗的成纤维细胞中的上调活性,药理学CK2抑制剂TBB(4,5,6,7-四氢苯甲酰苯二唑)显着消除了纤维细胞激活和氧化应激。 我们的数据提供了对心肌细胞对心脏成纤维细胞串扰的分子见解,以及CK2在调节心脏成纤维细胞激活和氧化应激中的潜在作用。激酶底物富集分析磷酸肽在此重塑过程中激酶(CK2,CDK2,PKC,GSK3B)的潜在作用。我们验证了酪蛋白激酶2(CK2)在分泌理论治疗的成纤维细胞中的上调活性,药理学CK2抑制剂TBB(4,5,6,7-四氢苯甲酰苯二唑)显着消除了纤维细胞激活和氧化应激。我们的数据提供了对心肌细胞对心脏成纤维细胞串扰的分子见解,以及CK2在调节心脏成纤维细胞激活和氧化应激中的潜在作用。

丹麦的量子相关网络安全
PKC(也称为非对称密钥加密)最初开发于 20 世纪 70 年代,通常与 RSA 同义,RSA 是第一个向公众开放的公钥密码系统。它解决了密码系统广泛部署的一个主要障碍:密钥交换。它使用公钥(共享)加密消息,使用私钥(秘密)解密消息。人们发现,通过利用解决一类称为单向函数的数学问题所涉及的计算难度,可以非常安全地做到这一点。例如,将两个非常大的素数相乘很容易,但从乘积中导出素因数却非常困难。当今部署的绝大多数公钥密码系统,包括 RSA 的后继者椭圆曲线密码系统 (ECC),都是基于这一单向函数原理。

非小细胞肺癌 MET 外显子 14 跳跃变异的当前和未来治疗选择
近膜 (JX) 结构域,其中包含 PKC 磷酸化位点 (S985)、胱天蛋白酶切割位点 (D1002) 和 E3 泛素连接酶 CBL (Casitas-B 系淋巴瘤) 对接位点 (Y1003),均控制 RTK 活性的下调 (图 1a)。3–7 这种改变破坏了外显子 14 两侧的内含子剪接位点,包括内含子 13 的剪接受体位点和内含子 14 的剪接供体位点,或外显子 14 编码序列本身内的突变,都会导致外显子 14 在转录本中跳跃。这些突变中最常见的是碱基替换,其次是插入/缺失。因此,导致MET外显子14跳跃的可变剪接事件会激活MET-HGF通路,促进肿瘤细胞增殖、迁移,并阻止细胞凋亡(图1b)。

早期糖尿病性视网膜病中神经退行性的细胞和分子机制
1得克萨斯大学里奥格兰德分校背景糖尿病性视网膜病(DR)仍然是美国人时代失明的主要原因。尚未有任何有效的治疗方法可以防止病情发作,只是治疗后期疾病。对疾病早期迹象的研究表明,视网膜神经层的变化是最早的疾病迹象,是在当前定义DR的血管变化之前。这引起了人们对DR涉及的神经变性的发病机理的兴趣。本综述解释了当前对DR中神经元变性的细胞和分子机制的理解,以及针对每种机制研究的潜在药理干预措施。方法进行了文献综述,以查看已定义并与DR相关的神经变性的每个主要细胞和分子途径,有关药理学干预措施的最新研究以及视网膜神经细胞与糖尿病中的微腔之间的关系,以促进神经变性。文章来自PubMed或最新的文章。结果多元醇,PKC,己胺和年龄途径已显示在高血糖中上调。多元途径描述NADPH,这是谷胱甘肽再生所必需的。神经细胞变得无法忍受ROS。果糖和山梨糖醇积聚在细胞中,导致肿胀。epalrestat,FDA批准糖尿病神经病以靶向醛糖还原酶,具有DR的潜力。PKC和rage途径促进了产生ROS的NADPH氧化酶。PKC-抑制剂Ruboxistaurin一直在临床试验中治疗糖尿病性视网膜病。己糖胺途径中间葡萄糖对线粒体有毒,并促进过氧化葡萄糖。benfotiamine,一种B1衍生物,可能会抑制年龄,PKC和六胺途径。dm会导致pro-nGF/ngf比率的不平衡,从而促进凋亡。NGF眼滴显示通过标准化比例来治疗DME的希望。BDNF比率也以相同的方式影响。持续补充BDNF会抑制光感受器的死亡,但是常规注射无效。DM发作后一周在视网膜组织中看到升高的TNF-升高,刺激外部凋亡。eTanercept,TNF-抑制剂,似乎会减慢DR的进展。高血糖下调用于神经元存活的PI3K/AKT途径。胰岛素促进了这种保护侵蚀凋亡的途径,但同时促进了凋亡。muller细胞和小胶质细胞被高血糖激活并释放炎症介质并引起谷氨酸兴奋性毒性。Muller细胞激活在DM发作后1.5个月,在6周内瞬时BBB分解以及胶质反应性提高。tau调节是由星形胶质细胞介导的。异常TAU引起星形胶质细胞功能障碍并导致神经元死亡。一生氧化物被ROS形成毛的硝酸盐并创造神经毒性环境而被灭活。VEGF促进了低水平的神经元存活,但通过高水平的BDNF和GNDF降解而凋亡。升高的ROS可促进VEGF并抑制其保护作用。结论已经描述了细胞和分子的糖尿病性视网膜血管病之前神经退行性的几种机制。许多研究详细介绍了导致视网膜血管病的神经退行性途径的潜力。继续研究哪种机制是开发有效治疗以防止DR发作的必要条件。

在计算机特征中
我们研究了短长核苷酸序列的硅硅表征,这些核苷酸序列在死亡应力诱导的转录组分析中差异表达。他们表现出与C末端旋转肽和防御素样蛋白的同源性,从而揭示了它们的抗菌活性。他们的预测纤维印刷显示出与抗菌肽有关的蛋白质特征。这些短长的RGA已显示具有结构性基序,例如APLT P型ATPase,酪蛋白激酶II(CK2),蛋白激酶3,蛋白激酶C(PKC)和N-糖基化位点,它们是抗病基因的属性。在配体对接分析中活跃结合位点精氨酸和赖氨酸残基的预测将它们作为抗菌肽预测,因为它们与抗菌活性的密切关系。硅结构 - 功能表征已经预测了它们在抗微生物病原体的抗性中的作用。此外,预测的抗菌肽区域显示了它们与Pr-5样蛋白和AMP家族Thaumatin

它比SSI/DID
报告处理协会主任•国际会议ASIACRYPT,CT-RSA,FC,PKC,ESORICS,ACNS,ASIACCS等。该计划主席。•FC指导委员会,FC 2013,2025执行(联合)主席•MyDataJapan General Informatored Association的副主席•可信赖的Web促进委员会TF的成员,可信赖的网络促进Web促进委员会委员会成员•数字机构DIW Advisory Board的成员DIW Advisory Board diw Advisory Group•使用数字公共工程型竞争•Financial Grodent Informity internage Interviation Intervient•使用数字公共工业工业工业委员会,贸易机构,贸易委员会,贸易委员会,贸易委员会,交易委员会,交易。关于如何回应数字和分散的金融•2016 - 2020年索夫林基金会董事会

确保在量子中确保在线安全
用于消息加密,仅受到已知量子计算攻击的部分影响。将当前加密密钥的长度加倍会减轻量子计算机的攻击。不对称的加密(例如RSA)是PKC的基础,并且依赖于困难的数学问题(分解质数是最常见的)。非对称加密图被广泛用于数字步骤中,例如签名和密钥交换,以确保对使用Internet至关重要的通信和网络。这些包括电子邮件交换,虚拟专用网络(VPN),安全网页连接,大多数形式的电子商务以及数字供应链等。对称和不对称的加密 - 策略通常在一起使用:不对称的加密式来执行关键的建立和一致,以及对https协议中的消息加密的对称性加密,最广泛用于Web浏览。