XiaoMi-AI文件搜索系统
World File Search SystemPhage

利用噬菌体展示技术改善肽的药代动力学
摘要:噬菌体展示是一种多功能方法,常用于发现针对疾病相关生物标志物的肽。该技术的主要优势在于亲和力选择(也称为生物淘选)的简便性和成本效益,可用于识别新肽。虽然识别具有最佳结合亲和力的肽相对简单,但所选肽的药代动力学通常被证明是次优的。因此,仔细考虑实验条件,包括选择使用体外、原位或体内亲和力选择,对于生成具有高亲和力和特异性并表现出理想药代动力学的肽至关重要。具体而言,体内生物淘选或体外、原位和体内亲和力选择的组合已被证明会影响肽和肽结合纳米粒子的生物分布和清除。此外,还必须考虑肽和纳米粒子之间性质的显著差异。虽然肽的生物分布主要取决于生理化学性质,并且可以通过氨基酸修饰进行改变,但纳米颗粒的大小和形状也会影响吸收和分布。因此,优化所需的药代动力学特性应是生物淘选策略中的一个重要考虑因素,以便能够选择有效靶向体内生物标志物的肽和肽结合纳米颗粒。

噬菌体 x 右侧操纵子的核苷序列
限制性片段。为了制备微克量的 Hin 375、Hin 550 和 Hae 790(见图 1),将含有示踪量 lambda [32p]_ DNA(2 X 106 cpm)的 5 mg 纯化 lambda DNA 用 Hin(7)或 Hae(6)消化,乙醇沉淀,重悬于 500 ul DNA 缓冲液(5 mM NaCi、10 mM Tris-HCl,pH 7.4、1 mM EDTA)中,在含有 TBE(1)缓冲液的 3.5% 聚丙烯酰胺凝胶(6 mm X 20 cm X 40 cm)上以 320 V 电泳 23 小时。通过放射自显影定位含有适当限制性片段的凝胶部分,切除,并通过苯酚提取去除 DNA(10)。如前所述,从含有 32P 的 DNA 中分离出高比活度标记的限制性片段(2)。通过聚丙烯酰胺凝胶电泳确定每个片段的链长(1、2)。
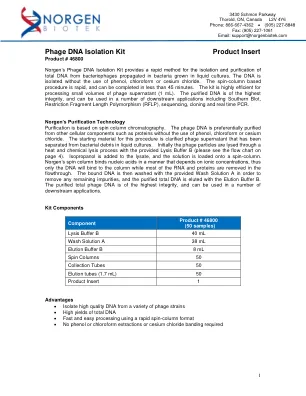
噬菌体 DNA 分离试剂盒产品说明书
噬菌体 DNA 分离试剂盒产品说明书 产品编号 46800 Norgen 的噬菌体 DNA 分离试剂盒提供了一种快速方法,可从在液体培养的细菌中繁殖的噬菌体中分离和纯化总 DNA。无需使用苯酚、氯仿或氯化铯即可分离 DNA。基于旋转柱的程序速度很快,可在 45 分钟内完成。该试剂盒可高效处理少量噬菌体上清液 (1 mL)。纯化的 DNA 具有最高的完整性,可用于多种下游应用,包括南方印迹、限制性片段长度多态性 (RFLP)、测序、克隆和实时 PCR。Norgen 的纯化技术 纯化基于旋转柱层析。无需使用苯酚、氯仿或氯化铯,即可优先从其他细胞成分(如蛋白质)中纯化噬菌体 DNA。该程序的起始材料是澄清的噬菌体上清液,该上清液已从液体培养物中的细菌碎片中分离出来。首先,使用提供的裂解缓冲液 B 通过热和化学裂解过程裂解噬菌体颗粒(请参阅第 4 页的流程图)。将异丙醇添加到裂解物中,然后将溶液加载到旋转柱上。Norgen 的旋转柱以取决于离子浓度的方式结合核酸,因此只有 DNA 会与柱结合,而大多数 RNA 和蛋白质会在流过中被去除。然后用提供的洗涤溶液 A 洗涤结合的 DNA 以去除任何残留杂质,并用洗脱缓冲液 B 洗脱纯化的总 DNA。纯化的总噬菌体 DNA 具有最高的完整性,可用于许多下游应用。试剂盒组件

使用
噬菌体是感染细菌并使用其宿主机械复制的病毒。它们是生物圈中最普遍,最多样化的生物学实体之一,具有较长的进化史。由于抗菌素耐药性(AMR)水平的增加和新发育的抗生素数量减少,人们对噬菌体治疗剂的兴趣更新。噬菌体在食品安全,水质,生物防治,疫苗和全球营养周期中也有应用。细菌和噬菌体都采用内部和外部自卫策略相互竞争,从而驱动基因组进化。虽然已经对许多噬菌体基因组进行了测序和注释,但噬菌体的蛋白质组学和脂肪组谱却几乎没有被探索,尤其是在感染阶段和溶酶体方面。本评论强调了在添加机器学习等工具的情况下,需要在-omics级别表征噬菌体 - 宿主关系。通过进一步了解噬菌体及其宿主之间的动态相互作用,可以利用合成生物学来设计新的解决方案,以应对我们当前的全球健康,农业和环境挑战。
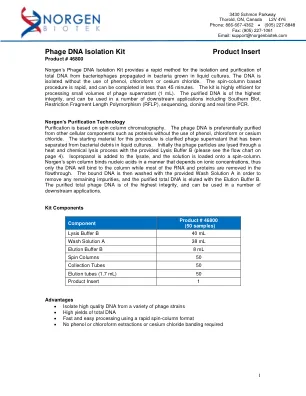
噬菌体 DNA 分离试剂盒产品说明书
噬菌体 DNA 分离试剂盒产品说明书 产品编号 46800 Norgen 的噬菌体 DNA 分离试剂盒提供了一种快速方法,可从在液体培养的细菌中繁殖的噬菌体中分离和纯化总 DNA。无需使用苯酚、氯仿或氯化铯即可分离 DNA。基于旋转柱的程序速度很快,可在 45 分钟内完成。该试剂盒可高效处理少量噬菌体上清液 (1 mL)。纯化的 DNA 具有最高的完整性,可用于多种下游应用,包括南方印迹、限制性片段长度多态性 (RFLP)、测序、克隆和实时 PCR。Norgen 的纯化技术 纯化基于旋转柱层析。无需使用苯酚、氯仿或氯化铯,即可优先从其他细胞成分(如蛋白质)中纯化噬菌体 DNA。该程序的起始材料是澄清的噬菌体上清液,该上清液已从液体培养物中的细菌碎片中分离出来。首先,使用提供的裂解缓冲液 B 通过热和化学裂解过程裂解噬菌体颗粒(请参阅第 4 页的流程图)。将异丙醇添加到裂解物中,然后将溶液加载到旋转柱上。Norgen 的旋转柱以取决于离子浓度的方式结合核酸,因此只有 DNA 会与柱结合,而大多数 RNA 和蛋白质会在流过中被去除。然后用提供的洗涤溶液 A 洗涤结合的 DNA 以去除任何残留杂质,并用洗脱缓冲液 B 洗脱纯化的总 DNA。纯化的总噬菌体 DNA 具有最高的完整性,可用于许多下游应用。试剂盒组件

RNA靶向CRISPR-Cas13提供广谱噬菌体免疫
。CC-BY-ND 4.0 国际许可下可用(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者此版本于 2022 年 3 月 26 日发布。;https://doi.org/10.1101/2022.03.25.485874 doi:bioRxiv 预印本

使用噬菌体显示以选择结构,折叠和...
折叠式和展开的分子选择用于热力学稳定性的选择是最新的发展是使用噬菌体显示器来选择具有改善热力学性能的蛋白质。通常,蛋白质稳定性是生物技术应用中的关键因素,无论是在升高温度还是在37°C下在生物医学应用中延长持续时间,并且通常与蛋白质搁板寿命相关。只有只有正确折叠的完整分子,因此功能结合位点才能与固定的配体相互作用,只要非本性蛋白质典型的非特异性相互作用可以有效地选择,则该形式可以通过噬菌体显示。在这些条件下,只要没有其他突变改变结合位点,功能性配体结合的选择有利于在噬菌体上更高的多肽突变体,即噬菌体,即较高百分比的分子位于本地状态的多肽突变体。作为一个序列,使用噬菌体显示的“正常”选择始终包括正确折叠的库成员的固有选择,因此在可接受的总体属性中选择了“复合”选择。有几位研究者[16-18]指出了这种观察结果,并在一项研究[19]中进行了系统的测试并证明,其中最佳折叠和最稳定的SCFV(单链抗体片段)可以从具有识别结合常数的一组SCFV中选择,但具有不同的热力学和折叠性和折叠性质。

长期认知活性期间的注意力脑节律
(未通过同行评审认证)是作者/资助者。保留所有权利。未经许可就不允许重复使用。此预印本版的版权持有人于2023年3月26日发布。 https://doi.org/10.1101/2023.03.03.26.534254 doi:biorxiv preprint
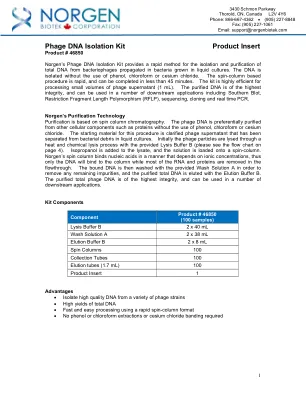
噬菌体DNA隔离套件产品插件
NORGEN的纯化技术纯化基于自旋色谱柱色谱法。噬菌体DNA优先纯化从其他细胞成分(例如蛋白质)中纯化,而无需使用苯酚,氯仿或氯化葡萄球菌。此过程的起始材料被阐明了噬菌体上清液,该噬菌体上清液已与液体培养物中的细菌碎片分离。最初,噬菌体颗粒通过提供的裂解缓冲液B通过热和化学裂解过程裂解(请参阅第4页的流程图)。异丙醇被添加到裂解物中,并将溶液加载到自旋柱上。Norgen的自旋柱以取决于离子浓度的方式结合核酸,因此只有DNA才能与柱结合,而大多数RNA和蛋白质在流潮中除去。然后用提供的洗涤溶液A洗涤结合的DNA,以去除剩余的杂质,并用洗脱缓冲液洗脱纯化的总DNA。纯化的总噬菌体DNA是最高的完整性,可用于许多下游应用。

美国的噬菌体疗法前进
噬菌体疗法是一种潜在的抗生素耐药性感染疗法,但在美国不像世界其他地区那样普遍使用。噬菌体疗法是前苏联和俄罗斯治疗细菌感染的历史实践。由于噬菌体自然存在于环境中,因此只有经过基因工程的合成噬菌体才能获得制药公司的专利,这使得在美国很难融入临床护理。但是,抗生素耐药性和生物技术的最新进展的日益增长的成本正在促使美国政府机构与工业合作,以支持开发合成噬菌体以打击抗生素耐药性。尽管很少有噬菌体疗法临床试验在过去的第二阶段进行了进展,但进一步发展的潜力令人难以置信。本综述通过评估噬菌体耐药性的风险与其潜在的益处,作为针对细菌耐药机制并提高抗生素易感性的有效产品,评估了美国噬菌体疗法的前景。