XiaoMi-AI文件搜索系统
World File Search SystemPosters




MER1173 ESMO海报(打印)
1。herpers b和al。(2022)NAT癌症3,418–436。2。Sching H和Al。 (2022)mol扬声器21,178。 3。 MJ Parsons和Al。 (2021)Discov 11,2413–2429。 4。 xu l,and al。 (2019) 5。 Katoh M.(2017)它们是J Oncol 51,1357–1 6。 cohen e和al。 (2023)癌症Res 83(补充8),CT012 [Oral,AACR 2023]。 7。 Fayew J和Al。 (2024)J Clin Oncol 42(补充16),摘要6014 [Oral,ASCO 2024]。 8。 sung h和al。 (2021)Ca Cinal 71,209–249。 9。 KJ的Harrington和Al。 (2023)J Clin Oncol 41,790–802。 10。 标记。 (2024)非正式处方。 可在以下网址提供:hwps://www.merck.com.com.com.com/usa/usa/usa/usa/kircurdas/kkey/kkey/eytruda/eye.pdf(2024年8月30日访问)。 11。 Wu Z和Al。 (2017)他们是J 12。 aj和al。 (2015)口服外科医学口腔Pathol 119,436–440。 13。 HK HOWON和AL。 (2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。Sching H和Al。(2022)mol扬声器21,178。3。MJ Parsons和Al。(2021)Discov 11,2413–2429。4。xu l,and al。(2019)5。Katoh M.(2017)它们是J Oncol 51,1357–1 6。 cohen e和al。 (2023)癌症Res 83(补充8),CT012 [Oral,AACR 2023]。 7。 Fayew J和Al。 (2024)J Clin Oncol 42(补充16),摘要6014 [Oral,ASCO 2024]。 8。 sung h和al。 (2021)Ca Cinal 71,209–249。 9。 KJ的Harrington和Al。 (2023)J Clin Oncol 41,790–802。 10。 标记。 (2024)非正式处方。 可在以下网址提供:hwps://www.merck.com.com.com.com/usa/usa/usa/usa/kircurdas/kkey/kkey/eytruda/eye.pdf(2024年8月30日访问)。 11。 Wu Z和Al。 (2017)他们是J 12。 aj和al。 (2015)口服外科医学口腔Pathol 119,436–440。 13。 HK HOWON和AL。 (2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。Katoh M.(2017)它们是J Oncol 51,1357–16。cohen e和al。(2023)癌症Res 83(补充8),CT012 [Oral,AACR 2023]。7。Fayew J和Al。 (2024)J Clin Oncol 42(补充16),摘要6014 [Oral,ASCO 2024]。 8。 sung h和al。 (2021)Ca Cinal 71,209–249。 9。 KJ的Harrington和Al。 (2023)J Clin Oncol 41,790–802。 10。 标记。 (2024)非正式处方。 可在以下网址提供:hwps://www.merck.com.com.com.com/usa/usa/usa/usa/kircurdas/kkey/kkey/eytruda/eye.pdf(2024年8月30日访问)。 11。 Wu Z和Al。 (2017)他们是J 12。 aj和al。 (2015)口服外科医学口腔Pathol 119,436–440。 13。 HK HOWON和AL。 (2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。Fayew J和Al。(2024)J Clin Oncol 42(补充16),摘要6014 [Oral,ASCO 2024]。8。sung h和al。(2021)Ca Cinal 71,209–249。9。KJ的Harrington和Al。 (2023)J Clin Oncol 41,790–802。 10。 标记。 (2024)非正式处方。 可在以下网址提供:hwps://www.merck.com.com.com.com/usa/usa/usa/usa/kircurdas/kkey/kkey/eytruda/eye.pdf(2024年8月30日访问)。 11。 Wu Z和Al。 (2017)他们是J 12。 aj和al。 (2015)口服外科医学口腔Pathol 119,436–440。 13。 HK HOWON和AL。 (2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。KJ的Harrington和Al。(2023)J Clin Oncol 41,790–802。10。标记。(2024)非正式处方。可在以下网址提供:hwps://www.merck.com.com.com.com/usa/usa/usa/usa/kircurdas/kkey/kkey/eytruda/eye.pdf(2024年8月30日访问)。11。Wu Z和Al。(2017)他们是J12。aj和al。(2015)口服外科医学口腔Pathol 119,436–440。13。HK HOWON和AL。 (2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。HK HOWON和AL。(2019)Expan Mol Med 51,1-1 14。 Xu MJ和Al。 (2017)Rev Mentastasis Cancer 36,463–473。(2019)Expan Mol Med 51,1-114。Xu MJ和Al。(2017)Rev Mentastasis Cancer 36,463–473。(2017)Rev Mentastasis Cancer 36,463–473。

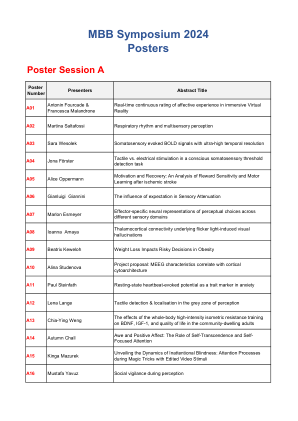

研讨会议程、摘要和海报
我们提供两个本科课程:生物科学学士学位和环境科学学士学位。生物科学课程仅对女性学生开放,为学生提供坚实的生物科学基础,使他们具备在新兴领域、市场和政策讨论中发挥领导作用的能力。环境科学课程有两个方向:海洋科学和生物技术,均获得 CHES(英国)认证。在研究生阶段,我们的环境科学理学硕士(也获得 CHES 认证)可以全日制或非全日制学习,而我们的生物和环境科学博士学位提供四个方向:细胞和分子生物学、环境科学、生态系统和海洋科学以及生物技术。此外,新的环境科学研究生证书旨在为学生和专业人士进一步的研究生学习做好准备,在达到绩效标准后,学生和专业人士有可能进入理学硕士课程的第二年学习。我们的部门寻求聪明、有上进心、勤奋的学生,他们渴望在专业或科学方面发挥领导作用,为自己的领域做出贡献,并为卡塔尔和卡塔尔大学服务。

doe fy 2022预算卷1
NNSA国防计划的战略伙伴关系计划办公室(NA-10.1)与联邦和非联邦实体建立并维持合作伙伴关系,以帮助维持美国的核威慑和更广泛的国家安全任务。战略合作计划办公室分为三个职能支柱:合作伙伴关系,机构计划和跨裁切支持。我的工作转移了所有三个组织活动。


MGFA 科学会议计划和海报
以下是已接受的 MGFA 科学会议海报,按主要作者的姓氏排列。MGFA 海报编号以“MG”开头,表示海报的位置(见下面的平面图)。还包括 AANEM 发布的《肌肉与神经》补充材料中列出的海报编号。