XiaoMi-AI文件搜索系统
World File Search SystemPrecise

Tow Assist如何支持主要设施的精确交付
元7建于东港院子。它配备了混合电力推进系统,该系统将柴油发动机与电池和双固定式方位角方位角推进器(US 255 S P30 fp)与电动感应电动机相结合。柴油发动机用于发电,然后将其存储在电池中。电动机可用于独立推动容器或与柴油发动机结合使用。在这种集成的功率和推进系统中,其他孔斯堡海事技术包括:•具有646KWH储能系统的K-Power DC电气系统•变压器•自动化和控制(K-Chief 700)•Shore Connection(用于充电)。
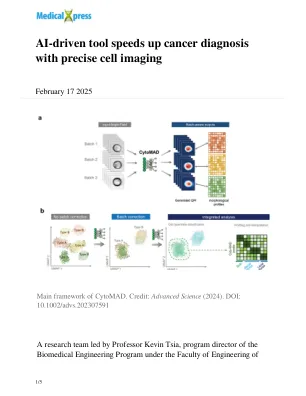
AI驱动的工具加快了精确细胞成像的癌症诊断
这种强大的深度学习模型受益于TSIA团队也开发的超快光学成像技术。“这项技术使我们能够以极高的速度捕获手机图像。每天都可以生成数千万的图像。因此,利用这一单个系统,我们处于许多AI创新中,我们处于一个独特的位置,以加速先进的AI R&D,从培训,优化到部署,”

在锂提取中精确离子筛分的聚酰胺膜互层的纳米形态发生
纳米过滤(NF)提供了一种可扩展且节能的方法,用于从盐湖中提取锂。然而,由于其水合离子半径的紧密相似性,锂与镁的选择性分离,尤其是在镁浓度高的盐水中,仍然是一个重大挑战。有限的LI + / mg 2 +当前NF膜的选择性主要归因于对孔径和表面电荷的控制不足。在这项研究中,我们报告了结合功能化的磺化carge胶以调节界面聚合过程的层间薄膜复合材料(ITFC)膜的发展。该集成的层间在控制胺基单体的扩散和空间分布中起着至关重要的作用,从而导致形成致密的纳米条纹聚酰胺网络。与常规的TFC膜相比,这些结构改进,包括精致的孔径和减少负电荷可显着提高LI + /Mg 2 +选择性(133.5)和渗透率增加2.5倍。此外,纳米条纹结构优化了膜过滤区域,同时最大程度地降低了离子传输抗性,从而有效克服了离子选择性和渗透性之间的传统权衡。这项研究强调了ITFC膜在达到高锂纯度和恢复的潜力,为大规模从盐水中提取大规模锂的途径有前途的途径。

利用逆转录衍生 DNA 对生命界进行精确的基因组编辑
外源 DNA 可以作为精确编辑细胞基因组的模板。然而,将体外产生的 DNA 输送到靶细胞可能效率低下,模板 DNA 的低丰度可能是精确编辑率低的原因。在细胞内产生模板 DNA 的一个潜在工具是逆转录子,这是一种参与噬菌体防御的细菌逆转录元件。然而,很少有人致力于优化逆转录子以产生设计序列。在这里,我们确定了逆转录子非编码 RNA 的修饰,这些修饰会产生更丰富的逆转录 DNA。通过测试能够实现高效逆转录的逆转录子操纵子架构,我们发现 DNA 产量的提高可以从原核细胞移植到真核细胞,从而实现更高效的基因组编辑。最后,我们表明逆转录子 RT-DNA 可用于精确编辑培养的人类细胞。这些实验为使用逆转录子产生 DNA 进行基因组修饰提供了一个通用框架。

植物引物编辑器实现水稻细胞的精准基因编辑
基因组编辑正在彻底改变植物研究和作物育种。序列特异性核酸酶 (SSN),例如锌指核酸酶 (ZFN) 和 TAL 效应核酸酶 (TALEN),已用于产生位点特异性 DNA 双链断裂并通过促进同源定向修复 (HDR) 实现精确的 DNA 修饰 (Steinert 等人,2016 年;Voytas,2013 年)。后来,RNA 引导的 SSN,例如 CRISPR-Cas9、Cas12a、Cas12b 及其变体,已应用于植物基因组编辑 (Li 等人,2013 年;Nekrasov 等人,2013 年;Tang 等人,2017 年;Zhong 等人,2019 年;Ming 等人,2020 年;Tang 等人,2019 年)。然而,HDR 依赖于 SSN 和 DNA 供体的同时递送,这在植物中一直具有挑战性( Steinert 等,2016; Zhang 等,2019)。在植物中实现高效 HDR 的另一个挑战是,在大多数细胞类型中,DNA 修复倾向于非同源末端连接(NHEJ)途径而不是 HDR( Puchta,2005; Qi 等,2013)。与受供体选择和 DNA 修复机制限制的 SSN 诱导的 HDR 不同,近年来开发的胞苷或腺嘌呤碱基编辑器可以在原型间隔物中 3-8 个核苷酸靶向窗口内将 C 转换为 T 或将 A 转换为 G( Komor 等,2016; Nishida 等,2016; Gaudelli 等,2017)。碱基编辑器虽然效率很高,但只能指导某些转换突变,而不能执行预定的颠换突变或插入和缺失 (indel)。在所有这些背景下,最近在人类细胞中开发所谓的引物编辑器 (PE) 方面取得的突破非常令人兴奋 ( Anzalone 等人,2019 )。在引物编辑中,Cas9H840A 切口酶与逆转录酶融合。融合蛋白在编辑 DNA 链上切口,通过引导到切口 DNA 并复制由引物编辑向导 RNA (pegRNA) 编码的遗传信息来启动逆转录。多功能的 pegRNA 是一种经过修饰的单向导 RNA (sgRNA),其 3' 端携带逆转录 (RT) 模板和引物结合位点 (PBS) 或序列中的引物。与 HDR 不同,PE 不需要 DNA 供体。在某些目标位点,PE 似乎也比碱基编辑器更精确、更高效(Anzalone 等人,2019 年)。

一种用于精确脑功能映射的启发式信息聚类搜索方法
图 3 (a) 与 ICS 算法相比,探照灯程序的步骤和输出的说明。灰色体素是探照灯方法中的搜索球中心体素,也是 ICS 算法中的起始体素。此示意图中探照灯的半径是一个体素,探照灯球的信息(每个球体用特定颜色表示)被分配给球体中心的体素,也就是说,输出图中的每个体素都具有与其搜索球相同的颜色(信息)。另一方面,ICS 方法的输出是一组通过数据驱动的启发式方法从起始体素扩展而来的聚类。每个聚类的信息都用特定的颜色表示。(b) 左:ICS 创建的重叠聚类的示例说明。右:单独描绘的相同聚类。黑点表示的体素是起始体素 vs,根据判别分析启发式方法进行扩展,从而为每个聚类得出特定的判别分数

透明,兼容的3D介质结构,用于精确评估器官的机械特征
最近开发了将薄膜材料的二维(2D)模式转换为3D介质结构的方法,在微系统设计中创造了许多有趣的机会。增长的感兴趣领域是多功能的热,电气,化学和光学接口到生物组织,尤其是3D多细胞,毫米尺度的构建体,例如球体,组装和类动物。本文提供了3D机械界面的示例,其中parylene-c的细丝带构成了透明,高度合规的框架的基础,这些框架可以可逆地打开和封闭,以捕获,包裹和机械限制脆弱的3D组织,以柔和的,非毁灭性的方式,以确切的粘膜属性测量,用于使用粘ellasticalsiques in nanoindent in nanoindentiques in nanoindentiques in nanoindentiques。有限元分析是一种设计工具,可用于指导对形状匹配的3D体系结构的几何和材料参数的选择。这些计算方法还量化了在打开和关闭其赋予的结构和力的过程中变形的各个方面,它们赋予了它们的结构和力。纳米识别的研究表明,根据器官的年龄,有效的Young的模量在1.5至2.5 kPa范围内。这一结果收集表明,在毫米级,软生物组织的非侵入性机械测量中广泛的效用。

简单的单体用于在开环的元置聚合过程中精确聚合物功能化
无-4.2 4.1 1.04 EXO-NI 2.0 4.7 4.9 4.9 1.02 ENDO-NI 1.9 4.7 5.3 1.02 ENDO-ONI 1.2 4.7 5.8 5.8 1.02 ENDO-ONI * ENDO-ONI * 1.0 4.5 5.0 1.02 ENDO-PONI 〜0 4.9 4.9 6.4 1.08使用1 H NMR SpectRoscopy计算了计算。b根据单体和催化剂的进料进行计算,并假设每个步骤都完全转换。c由THF中的三重检测尺寸排除色谱(SEC)确定,用狭窄的PMMA标准校准。

在不确定的环境接触和力量中,软机器人的精确控制
摘要 - 重新研究已经报道了生物启发的软机器人的显着性,以表现出灵敏和接触式的友好型。在这项工作中,我们采取了第一个步骤,通过提出一个全面的建模和控制框架来解决细长气动软机器人的问题。我们的框架采用了一个完全参数化的模型,该模型可以准确地描述了使用Hermite插值的机器人配置和分布力。利用此模型,我们进一步建立了一种估计算法,该算法可以推断出有限的运动数据中的完整机器人配置并分布外力,从而使接触位置和力量感知。整合了该模型和估计器,我们的控制框架 - 工作在不同的力下实现了精确的机器人运动控制,平均轨迹跟踪误差在0.3 mm之内。它还检测到并适应不确定的接触,在自动避免障碍物和精确抓握的测试中证明了这一点。此框架对各种应用程序(例如环境探索和安全操纵)有望在需要与环境的互动中进行安全操作。

斑马鱼中发育信号传导和人类病理的体内研究的精确基础编辑
摘要虽然斑马鱼正在成为研究人类疾病的新模型系统,但仍缺乏一种有效的方法来产生高效率的精确点突变。在这里,我们表明基本编辑者可以在没有其他不必要的靶向突变的情况下生成具有高效率的C-T点突变。此外,我们还建立了一个新的编辑变体,以识别NAA原始探针相邻基序,从而扩大了斑马鱼中的基本编辑可能性。使用这些方法,我们首先在CTNNB1基因中产生了基本变化,模仿了已知的人类基因的突变,从而导致内源性Wnt信号传导的组成型激活。此外,我们精确地针对了包括癌症相关的几个基因,包括CBL。使用了最后一个目标,我们创建了一个新的斑马鱼矮人模型。一起,我们的发现扩大了斑马鱼作为模型系统的潜力,允许新的方法调节细胞信号通路和人类遗传疾病相关突变的精确模型的生成。