XiaoMi-AI文件搜索系统
World File Search SystemUbiquitination

通过翻译后修改对胰岛素分泌的调节
胰岛素是一种从胰岛胰岛细胞中释放出的合成代谢激素,具有保持血糖稳态的独特能力(Cabrera等,2006; Da Silva Xavier,2018)。胰岛素对葡萄糖和其他营养素(例如氨基酸和游离脂肪酸)的分泌是一个复杂的过程,涉及多个信号通路的配位(Nolan等,2006; Henquin,2011)。这一过程的损害与糖尿病的发展直接相关(Schwartz等,2013)。因此,胰岛素分泌过程被认为是治疗糖尿病的有希望的靶标(Defronzo等,2014)。但是胰岛素分泌法规的细节仍然是一个未解决的问题。翻译后修饰(PTM)是在蛋白质上添加或去除化学基团的共价修改(Walsh等,2005)。通过调节蛋白质定位,降解和功能,它几乎与所有生理和病理过程密切相关(Walsh和Jefferis,2006; Khan等,2016; Morales-Tarre等,2021; Zhu and Hart and Hart,2021)。积累的证据表明,PTM广泛参与胰岛素分泌过程,目前,至少八种类型的PTMS已知与胰岛素分泌有关。例如,信号级联介导胰岛素分泌需要磷酸化(Campbell和Newgard,2021年)。sumoylation和棕榈酰化可以在多个阶段调节胰岛素分泌(Davey等,2019; Chamberlain等,2021)。这些表明乙酰化,泛素化和O-Glcnacylation与胰岛素基因转录有关(Mounier and Posner,2006; Ozcan等,2010; Sampley和Ozcan,2012)。甚至最近据报道,即使是一些研究的PTM,例如柠檬化和脱氨酸,也与胰岛素分泌有关。

肿瘤细胞中 MARCHF3 降解 PARP1 引发树突状细胞中 cCAS-STING 激活,从而调节肝细胞癌中的抗肿瘤免疫
摘要 背景 对免疫检查点抑制剂 (ICI) 的耐药性显著限制了肝细胞癌 (HCC) 患者免疫治疗的疗效。然而,免疫治疗耐药性的机制仍然不太清楚。我们的目的是在抗程序性细胞死亡蛋白 1 (PD-1) 治疗框架内阐明膜相关环 CH 型指 3 (MARCHF3) 在 HCC 中的作用。 方法 在对 ICI 表现出不同反应的 HCC 肿瘤的转录组谱中鉴定出 MARCHF3。在人类中,通过多重免疫组织化学评估 MARCHF3 表达与肿瘤微环境 (TME) 之间的相关性。此外,通过流式细胞术评估了肿瘤细胞中的 MARCHF3 表达和免疫细胞浸润。 结果 在对 ICI 有反应的患者的肿瘤中,MARCHF3 显著上调。HCC 细胞中 MARCHF3 表达的增加促进了树突状细胞 (DC) 成熟并刺激 CD8 + T 细胞活化,从而增强了肿瘤控制。从机制上看,我们确定 MARCHF3 是 DNA 损伤反应的关键调节因子。它通过 K48 连接的泛素化直接与聚(ADP-核糖)聚合酶 1 (PARP1) 相互作用,导致 PARP1 降解。该过程促进双链 DNA 的释放并激活 DC 中的 cCAS-STING,从而启动 DC 介导的抗原交叉呈递和 CD8 + T 细胞活化。此外,ATF4 转录调控 MARCHF3 表达。值得注意的是,PARP1 抑制剂奥拉帕尼增强了抗 PD-1 免疫疗法在皮下和原位 HCC 小鼠模型中的疗效。结论 MARCHF3 已成为 HCC TME 中免疫景观的关键调节因子,并且是 HCC 的有力预测生物标志物。将针对 DNA 损伤反应的干预措施与 ICI 相结合是一种有前途的 HCC 治疗策略。

1 HAO-FOUNTAIN综合征蛋白USP7通过新型p53独立的泛素3信号通路4 5
摘要35蛋白质泛素化的精确控制对于大脑发育至关重要,因此,泛素信号网络的破坏36可能导致神经系统疾病。37个去泛素酶USP7的突变导致HAO-Fountain综合征(HAFOUS),其特征是38个发育延迟,智力残疾,自闭症和侵略性行为。在这里,我们报告了39个小鼠前脑中兴奋性神经元中USP7的条件缺失触发了40种表型,包括感觉运动缺陷,学习和记忆力障碍以及侵略性的41个行为,类似于Hafous的临床特征。USP7缺失诱导神经元细胞凋亡的42依赖性肿瘤抑制剂p53。然而,尽管损失了p53,但43个USP7条件小鼠的大多数行为异常仍然存在。引人注目的是,大脑中的USP7缺失44突触蛋白质组和树突状脊柱形态发生独立于p53。综合45蛋白质组学分析表明,神经元USP7相互作用富含与神经发育疾病有关的蛋白质46,并专门鉴定了RNA剪接因子47 PPIL4作为USP7的新型神经元底物。皮质神经元中PPIL4的敲低会损害48个树突状棘的形态发生,表现USP7损失对树突状棘的影响。49这些发现揭示了一种新型的USP7-PPIL4泛素信号传导链接,该联系调节发育中的大脑中的神经元50连通性,这对我们对Hafous和其他神经发育障碍的发病机理51的理解产生了影响。52 53关键字54泛素,去泛素酶,USP7,HAO-Fountain综合征,p53,脑发育,55谷氨酸能神经元,突触,TMT蛋白质组学,PPIL4 56 56 57 58 59 59

theranostics USP14调节类似茎状的特性,...
原理:胶质母细胞瘤(GBM)是最具侵略性的原发性脑癌类型,并包含有助于肿瘤生长和治疗性抗性的自我更新GBM干细胞(GSC)。然而,对GSC治疗耐药性的分子决定因素知之甚少。方法:我们对患者衍生的GSC中的去泛素化酶(DUB)进行了全基因组分析,并使用基因特异性shRNA来识别有助于GSC存活和放射线抗性的重要DUB基因。随后,我们采用质谱和免疫沉淀来显示USP14和AlkBH5之间的相互作用,并确定了上游激酶MST4,这对于碱性化和稳定碱的稳定至关重要。此外,我们进行了集成的转录组和M 6 A-SEQ分析,以发现影响GSC辐射势的ALKBH5的关键下游途径。结果:我们的研究证明了去泛素酶USP14在维持GSC的干性,致癌潜力和放射线的重要作用。USP14通过防止其K48连接的泛素化和通过HECW2降解M 6 A脱甲基碱ALKBH5。通过MST4在丝氨酸64和69处的AlkBH5磷酸化增加了其与USP14的相互作用,从而促进了AlkBH5的去泛素化。此外,ALKBH5以取决于YTHDF2的方式直接与USP14转录本相互作用,建立了一个正反馈环,该反馈环维持GSC中两种蛋白质的过表达。暴露于电离辐射(IR)后,在GSC中进一步刺激了此信号级联。MST4-USP14-AlkBH5信号通路对于增强干细胞样性状,促进DNA双链断裂的同源重组修复以及促进GSC中的放射性和肿瘤性。用小分子IU1抑制USP14会破坏ALKBH5去偶联性,并提高IR疗法对GSC衍生的脑肿瘤异种移植物的有效性。结论:我们的结果将MST4-USP14-AlkBH5信号通路确定为治疗GBM的有前途的治疗靶标。

2021; 17(2): 417-429。doi: 10.7150/ijbs.54194 研究论文 SOX13/TRIM11/YAP 轴促进 anapla 的增殖、迁移和化学耐药性
间变性甲状腺癌 (ATC) 是最具侵袭性和恶性程度的实体肿瘤之一。泛素蛋白酶体系统存在于所有真核细胞中,对细胞稳态至关重要。但其在 ATC 中的潜在作用仍不清楚。TRIM11 是一种 E3 泛素连接酶,据报道在多种人类癌症中充当致癌基因。本研究旨在揭示 TRIM11 在 ATC 中的致癌功能。使用 Western blot 测量 TRIM11 和 YAP 的蛋白表达,而使用实时 PCR 测量 YAP 靶基因。CCK8 测定用于检测细胞活力;划痕愈合试验和 transwell 测定用于测量 ATC 的迁移能力。异种移植瘤模型用于体内研究。免疫沉淀试验用于检测 YAP 和 TRIM11 之间的相互作用域。并用基于泛素的免疫沉淀实验检测YAP发生的具体泛素化方式。TRIM11的缺失显著降低了ATC细胞的增殖和迁移能力,增加了细胞对化疗的敏感性,而YAP的过表达可以进一步挽救这种效应。TRIM11的缺失降低了ATC中YAP蛋白水平和YAP/TEAD靶基因,如CTGF、ANKRD1和CYR61。这表明TRIM11是Hippo信号通路的调控因子。免疫沉淀实验表明TRIM11的RING结构域对于与YAP的WW结构域的相互作用至关重要。进一步的机制分析表明TRIM11促进YAP的单泛素化,从而延长其蛋白半衰期。此外,TRIM11启动子分析表明SOX13通过与TRIM11启动子结合来激活TRIM11的转录。综上所述,本研究揭示了TRIM11在ATC中的致癌功能,TRIM11是Hippo通路的翻译后调控因子,靶向TRIM11可能是治疗ATC的一种潜在方法。
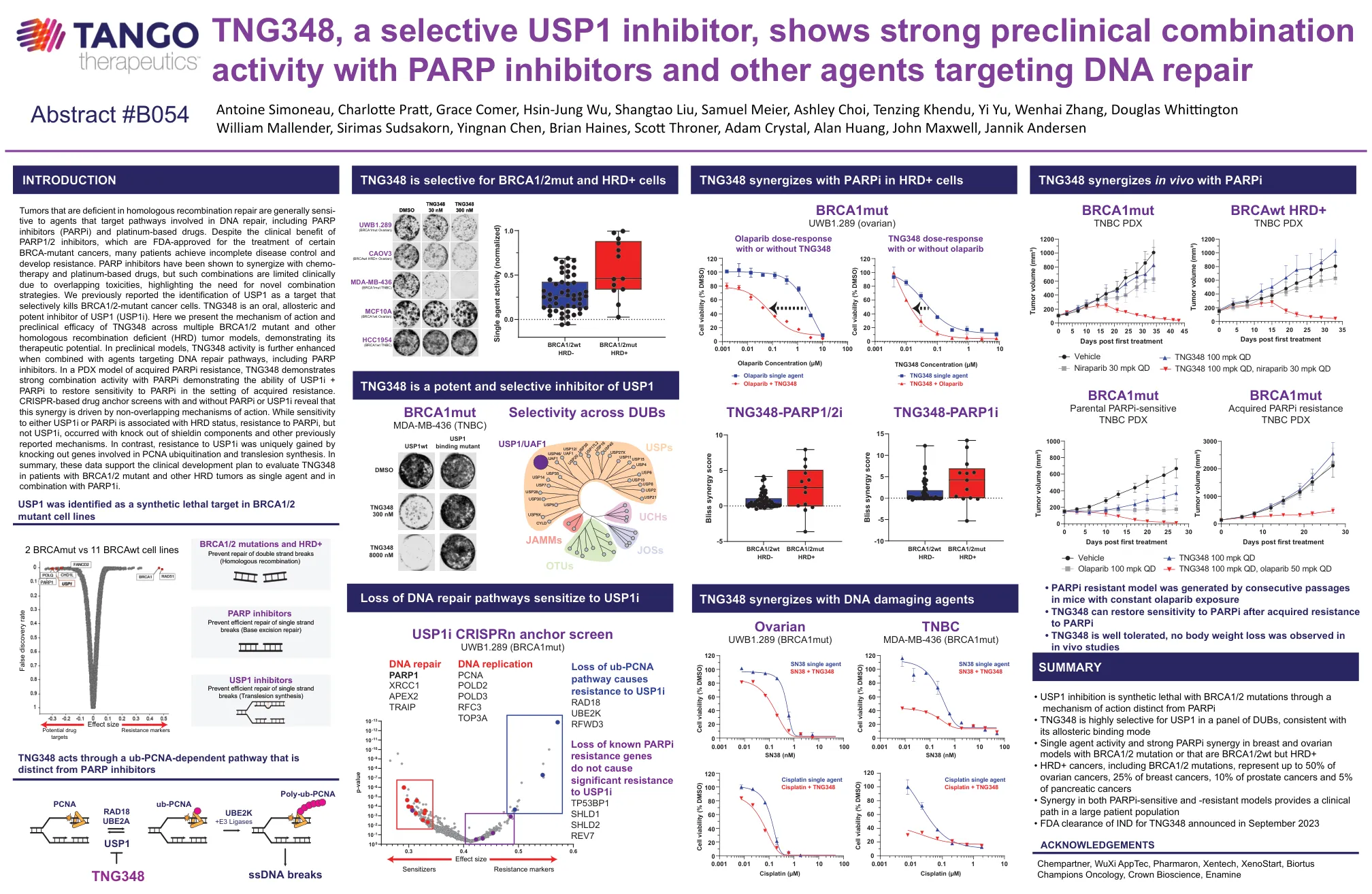
TNG348 海报 ENA 2023Final
同源重组修复缺陷的肿瘤通常对针对 DNA 修复途径的药物敏感,包括 PARP 抑制剂 (PARPi) 和铂类药物。尽管 PARP1/2 抑制剂具有临床益处,且已获 FDA 批准用于治疗某些 BRCA 突变癌症,但许多患者的疾病控制并不完全并且产生了耐药性。PARP 抑制剂已被证明可与化疗和铂类药物产生协同作用,但由于毒性重叠,此类组合在临床上受到限制,这凸显了对新组合策略的需求。我们之前报告已鉴定出 USP1 可选择性杀死 BRCA1/2 突变癌细胞的靶点。TNG348 是一种口服、变构和强效的 USP1 (USP1i) 抑制剂。这里我们介绍了 TNG348 在多种 BRCA1/2 突变体和其他同源重组缺陷 (HRD) 肿瘤模型中的作用机制和临床前疗效,证明了其治疗潜力。在临床前模型中,当与针对 DNA 修复途径的药物(包括 PARP 抑制剂)结合时,TNG348 活性会进一步增强。在获得性 PARPi 耐药性的 PDX 模型中,TNG348 表现出与 PARPi 的强大组合活性,证明了 USP1i + PARPi 能够在获得性耐药的情况下恢复对 PARPi 的敏感性。有和没有 PARPi 或 USP1i 的基于 CRISPR 的药物锚定筛选表明,这种协同作用是由不重叠的作用机制驱动的。虽然对 USP1i 或 PARPi 的敏感性与 HRD 状态有关,但对 PARPi 而非 USP1i 的耐药性发生在屏蔽蛋白成分的敲除和其他先前报道的机制下。相比之下,通过敲除参与 PCNA 泛素化和跨损伤合成的基因,可以独特地获得对 USP1i 的抗性。总之,这些数据支持临床开发计划,以评估 TNG348 作为单一药物和与 PARP1i 联合用于 BRCA1/2 突变体和其他 HRD 肿瘤患者。

免疫疗法和癌症:多摩学观点
免疫疗法(IT)代表了癌症治疗方面的显着成就[1]。肿瘤免疫疗法通过重新启动肿瘤免疫周期并恢复人体的天然抗肿瘤免疫反应来起作用[2]。目前,至少有四种主要的免疫疗法策略,其中包括免疫检查点抑制剂(ICIS),例如程序性细胞死亡蛋白1(PD-1)和细胞毒性T淋巴细胞抗原4(CTLA-4),嵌合抗原受体T-Cell受体T-Cell Therof actapy,Tumory pacocine and Tumory pacocines,thmory和Peripications and Peripatications和Peripaticationcation。尽管这些疗法已广泛成功,但增强了临床肿瘤结局[2],但并非所有患者都从中受益[1]。因此,对于从免疫疗法中获得最多的筛查至关重要[2]。肿瘤异质性可能是由于遗传,表观遗传和转录修饰等多种因素而产生较低治疗疗效的原因。蛋白质表达变化;以及代谢谱的变化[3]。最近,人们非常关注翻译后修饰(PTMS),这些变化是对单个氨基酸的小变化,例如糖基化,乙酰化,乙酰化,磷酸化,棕榈酰化和泛素化或泛素化或去泛素化。已经发现这些PTM具有改变蛋白质与其他分子的功能,形状,平衡和相互作用的能力。此外,最近的研究表明,PD-1和程序性细胞死亡配体1(PD-L1)的表达水平可以受到表观遗传,转录和转录后系统的调节,从而影响肿瘤免疫[4,5]。在这种情况下,多词的方法结合了基因组学,转录组学,蛋白质组学,代谢组学,放射组学和免疫学,有助于揭示肿瘤中存在的各种层次,并探索蛋白质内的双重性,并探索蛋白质丰富的蛋白质,代表性地表现出跨性别的细胞表达,摩尔纳的形式和基因型的摩擦性,基因构图,基因构想,基因构图,基因范围,基因范围,莫尔纳(MRNA)的概述,莫尔纳(MRNA)的概述,莫尔娜癌症以及肿瘤 - 免疫间隔机制,鉴定出新的潜在生物标志物和免疫疗法靶标,并促进与免疫疗法相关的独特分子特征的鉴定

FUCCI实时细胞周期成像作为设计改进的癌症治疗的指南:针对靶向静态化学抗性癌细胞的创新策略的回顾
摘要:固体癌症化疗的进展在很大程度上是由于肿瘤中静态癌细胞的化学抗性而在悲惨的缓慢上缓慢。Miyawaki等人于2008年开发了荧光泛素化细胞周期指标(FUCCI),该指标是实时代码颜色的细胞周期的颜色。fucci利用与不同的颜色荧光报告基因相关的基因,这些基因仅在细胞周期的特定阶段表达,从而实时可以对细胞周期的阶段进行图像。肿瘤内经液的实时FUCCI成像表明,已建立的肿瘤包括大多数静态癌细胞和位于肿瘤表面的次要循环癌细胞,或靠近肿瘤血管。与大多数循环癌细胞相比,静止的癌细胞对细胞毒性化疗具有抗性,其中大多数靶细胞在S / G 2 / M期中。静止的癌细胞可以在存活治疗后重新进入细胞周期,这表明了大多数细胞毒性化学疗法通常对固体癌症有效的原因。因此,静止的癌细胞是癌症治疗的主要障碍。FUCCI成像可用于靶向肿瘤中静止的癌细胞。For example, we review how FUCCI imaging can help to identify cell-cycle-specific therapeutics that comprise decoy of quiescent cancer cells from G 1 phase to cycling phases, trapping the cancer cells in S / G 2 phase where cancer cells are mostly sensitive to cytotoxic chemotherapy and eradicating the cancer cells with cytotoxic chemotherapy most active against S / G 2 phase cells.fucci可以在实时的体外和体内轻松地在单细胞水平上图像细胞周期动力学。因此,可视化使用FUCCI肿瘤内的细胞周期动力学可以为许多改善固体癌症细胞周期靶向疗法的策略提供指南。

NSUN2 介导的 m5C 修改驱动替代......
原理:间变性甲状腺癌 (ATC) 是一种极具侵袭性的甲状腺癌,在初次诊断时经常表现为局部晚期浸润或远处转移,因此错过了手术干预的最佳窗口。因此,全身化疗和靶向治疗对于改善 ATC 的预后至关重要。然而,ATC 对常规治疗表现出显著的耐药性,这凸显了阐明这种耐药性背后的生物学机制并确定新的治疗靶点以克服它的必要性。方法:我们对来自 ATC 样本的大量和单细胞 RNA 测序 (scRNA-seq) 数据进行了全面分析,以筛选与多药耐药 (MDR) 相关的 m 5 C 修饰相关基因。然后,我们进行了 IC 50 测定、流式细胞术,并使用了 Nsun2 敲除的自发致瘤 ATC 小鼠模型来证明 NSUN2 促进了 ATC 中的 MDR。为了研究 NSUN2 介导的耐药机制,我们生成了 NSUN2 敲除的 ATC 细胞系并进行了转录组学、蛋白质组学和 MeRIP-seq 分析。此外,还进行了 RNA 测序和可变剪接分析以确定 NSUN2 敲除后的整体变化。我们通过糖蛋白染色、变性 IP 泛素化、核质分馏和 PCR 进一步探索了 NSUN2/SRSF6/UAP1 轴的潜在机制。最后,我们在体外和体内评估了小分子 NSUN2 抑制剂与抗癌药物的协同作用。结果:我们的研究结果表明,NSUN2 表达与 ATC 中的 MDR 显着相关。 NSUN2 充当 SRSF6 mRNA 上的 m 5 C 的“写入器”,ALYREF 充当 m 5 C 的“读取器”,从而诱导选择性剪接重编程并将 UAP1 基因的剪接形式从 AGX1 重定向到 AGX2。因此,AGX2 增强了 ABC 转运蛋白的 N 连接糖基化,通过防止泛素化介导的降解来稳定它们。此外,NSUN2 抑制剂可降低 NSUN2 酶活性并减少下游靶标表达,从而为克服 ATC 中的 MDR 提供了一种新颖且有希望的治疗方法。结论:这些发现表明 NSUN2/SRSF6/UAP1 信号轴在 ATC 的 MDR 中起着至关重要的作用,并将 NSUN2 确定为 ATC 化疗和靶向治疗的协同靶点。

遗传背景可以调节饲料添加剂的影响吗?肠道微生物组和转录组相互作用的答案,饲喂植物剂,有机酸或益生菌的混合物
尽管有可持续性,但在养殖鱼类中,选择性育种和饲料添加剂之间的协同作用仍然不足。参考(Ref)和选定的吉尔特黑头海bream生长(GS)在14天内用对照(CTRL)饮食喂食。ctrl饮食与三种功能添加剂(基于大蒜和中链脂肪酸的PHY:植物生成型; OA:有机酸混合物与70%的丁酸丁酸钠盐;概率:基于益生菌的有机酸混合物,益生菌,基于枯草菌,枯草脂,脓疱和licheniformes)。然后将这些实验饮食依次以高(PHY/OA = 7.5 g/kg,prob = 2×10 11 CFU/kg; 2周)和低(PHY = 5 g/kg,OA = 3 g/kg,prob = 3 g/kg,prob = 4×10 10 CFU/kg; 10 cfu/kg; 10周)。给定基因型和添加剂的能力来改变鱼类生长的性能,肠道健康以及宿主与其前肠(AI)微生物植物的相互作用。gs鱼显示出更好的生长和饲料转化率,与肠道微生物组成的个体变异性降低有关。PHY添加剂对GS-Phy鱼的肠道转录组有重大影响,并在上皮完整性,鞘脂和胆固醇/胆汁/胆汁盐代谢的上调上调。随着OA添加剂的增长性能,AI杯状细胞区域减少和AI粒细胞浸润的增强与中性粒细胞脱粒标记物的下调相关,与致病属的下降有关发酵和维生素K生物合成推断的途径。杆菌的建立和缺乏AI炎症在两个遗传背景的概率中平行。但是,GS鱼的生长和使用添加剂的饲料越来越好,而Ref Fish中出现了恶化。这种改善与硝酸盐还原kocuria的丰度,上皮细胞维持和增殖的标记的上调以及微生物群可调的蛋白质先素质和泛素化标记的下调有关,支持了上皮的较低的转离和改善的肠道范围。总的来说,吉尔特黑德海bream中营养创新的成功在很大程度上取决于宿主基因组易感性,也取决于肠道菌群cording to to Hologenome理论。