XiaoMi-AI文件搜索系统
World File Search SystemWADA

兽医学委员会
“Wood 博士表示,500 到 6,000 美元之间的金额可能比较合适。10,000 美元太高,而且要判处五年监禁。她认为众议院的法案过于激进。此外,第 5 页还谈到了兽医执行此程序时通常采用的剪尾或断尾方法。再说一次,她对剪尾断尾没有意见,但认为断尾断尾需要由有执照的兽医来做。”由于会议记录没有进一步的更正,经 Nishimoto 先生提出动议,Wada 女士附议,会议表决并一致通过,批准了 2023 年 12 月 8 日会议和 2024 年 2 月 2 日会议的修订会议记录。第 91 章,HRS,主席询问是否有任何公众成员愿意就此议程项目提供裁决口头证词。事项:Ivy Kim 律师举手并被提升为小组成员。 Kim 女士向委员会进行了自我介绍,她来自 DCCA 的 RICO 部门。她正在为 Malcome E. Hickman 的兽医执业执照问题第 2b 项提供证词。她想重申此案的进展情况。在本案中,RICO 提交了一封信和一份宣誓书供委员会审议。在本案中,和解是在去年 6 月达成并获得委员会批准的。自委员会批准和解协议以来,已经过去了近 9 个月。他已经支付了罚款,但他还没有完成要求他完成的 20 小时 CE。她了解到 Hickman 博士声称发生了计算机崩溃、技术故障,这也是他在 11 月给我们的同样借口。现在是 2024 年 3 月,我们仍然没有收到任何证明他完成了所需 CE 小时的证据。DAG 询问 Kim 女士是否通知了持照人?Kim 女士回答是的。她通过挂号信将宣誓书和证物寄给了希克曼博士,还通知他将于 2024 年 3 月 13 日举行听证会,并给了他董事会会议的日程安排和网站地址,邮件于 2024 年 2 月 6 日送达。DAG 询问金女士是否有 2024 年 2 月 6 日的送达回执?金女士说她确实有送达回执,看起来不是他的名字,而是别人领取了邮件。挂号信的回执上没有注明送达日期,但她有一个跟踪号,当她检查跟踪号时,确实显示邮件是在 2024 年 2 月 6 日送达的。

irt2024
核酸的研究合成第6节(主席:Kathie Seley-Radtk E)9:35-9:50 OP4 - Malgorzata Honcharenko,Karolinska Institutet一种新颖的方法,是一种合成寡核苷酸多核苷酸多核苷酸的新方法Peyrottes,蒙彼利埃大学,CNRS碳碳和氯核苷磷酸类似物作为恶性疟原虫抑制疟原虫的新型化学型10:15-10:30 OP6 - Robert Britton,Robert Britton,Simon Fraser University,快速,灵活,柔性,可稳定的,可伸缩的核心合成,the tea coffee teacoy 10:30:30:30:55 55 55 55 55 55(55 55) Asanuma)10:55-11:20 IL8 - 塞奇·范·卡伦伯格(Serge van Calenbergh),根特大学结核素类似物与重要的Human and Fivestock疾病相似的原生动物病原体11:25-11:40 OP7 - Nicholas Chim,Nicholas Chim,加利福尼亚大学,最大程度地融合了最有效的TRYMASE TRYMASE TRYMASE TRYMASE TRYMASE TRIMPSINGS TRIMPTIONT, 11:45-12:00 OP8 - Michal Hocek,捷克科学学院酶合成基础改性RNA与工程DNA聚合酶基础修饰的RNA 12:00-13:30午餐,海报II II次,第8届海报(主席:FUMI NAGATSUGI:FUMI NAGATSUGI)13:30-13:30-13:55 IL9 - ROGERSTRASES基于Rogerstrified Artrins on Artnified Artnifirent on strutt on strutt on strutt rocority rogation intriptiation寡核苷酸ES 14:00-14:15 OP9 - 加利福尼亚大学的Dong Wang,圣地亚哥分校的结构基础,是通过Cel-lular RNA聚合酶14:20-14:20-14:35 OP10-Michiko Kimoto,Xenolis Pte的转录遗传字母识别的遗传字母。Ltd. Six-Letter DNA Aptamer Generation as an Antibody Alternative 14:40-16:00 Coffee, tea Recruitment/Discussion session Session 9 (Chair: Ramon Erit ja) 16:00-16:25 IL10 – Kazuo Nagasawa, Tokyo University of Agriculture and Technology Control of functions of dynamically formed high-order nucleic acids by polyoxazole compounds 16:30-16:45 OP11 – M. Carmen Galan, University of Bristol Small molecule G-quadruplex ligands are antibacterial candidates for Gram- nega- tive bacteria 16:50-17:05 OP12 – Shigeori Takenaka, Kyushu Institute of Technology Double-strand structuring of oligo-thymine by cyclic bis-naphthalene diimide Session 10 (主席:Takehiko Wada)17:10-17:25 OP13 - Vyacheslav V. Massey University University结构的结构引导抑制癌症DNA-Mutating酶Apobec 3A 17:30-17:30-17:55 IL11 - Zlatko Janeba,Iocb purague pare

来自单个大脑部分的空间代谢组脂肪组和糖果
gentry 1,2, *,李陈3, *和拉蒙·C·太阳1,2, * 1 1佛罗里达州佛罗里达州盖恩斯维尔大学医学院生物化学与分子生物学系,美国佛罗里达州盖恩斯维尔大学2佛罗里达大学神经科学系,美国佛罗里达州盖恩斯维尔大学5成瘾研究与教育中心,佛罗里达大学,佛罗里达州盖恩斯维尔,佛罗里达州盖恩斯维尔6麦克奈特脑研究所,佛罗里达大学,佛罗里达州盖恩斯维尔大学,佛罗里达州7 7美国佛罗里达州盖恩斯维尔市9佛罗里达大学化学系,美国佛罗里达州盖恩斯维尔大学10年老化学院,佛罗里达大学,佛罗里达州盖恩斯维尔,美国#这些作者同等贡献:Harrison A. Clarke; Xin MA; Cameron J. Shedlock *这些作者共同监督这项工作:Matthew S. Gentry;李陈拉蒙·C·太阳摘要:代谢产物,脂质和聚糖是参与复杂生物系统的基本生物分子。它们通过定义生物体的生理学和病理学的无数途径和分子过程进行代谢引导。在这里,我们提出了一种蓝图,用于使用质谱成像从单个组织中对空间代谢组,脂肪组和糖的同时分析。个人赞美原始的实验协议,我们的工作流程包括一个称为空间增强多组学界面(SAMI)的计算框架,该框架提供了多组学的整合,高尺寸聚类,空间解剖学映射,具有匹配的多组学特征,以及为无效的互联网分配和互动的互动式分配,并提供匹配的多组学特征,并提供互动生物学。INTRODUCTION Metabolomics (Fiehn, 2002; Gibney et al., 2005; Lisec et al., 2006), lipidomics (Cajka and Fiehn, 2016; Han and Gross, 2005), and glycomics (Cummings and Pierce, 2014; Ruhaak et al., 2010; Wada et al., 2007) are three distinct facets of omics methodologies, each offering a unique window进入活生物体中相连且复杂的生化过程。这些领域的当前状态缺乏空间分辨率和统一的综合分析,这些分析提供了互连代谢景观的广泛概述。发展空间分辨的代谢组学,脂质组学和糖基因对于促进我们对生物系统的了解至关重要,并且有可能改变我们对复杂组织代谢异质性的理解,发现新型的生物标志物甚至治疗靶标。然而,这种综合方法的发展受到每个分子类别的理化特性和分析要求的固有差异的挑战。基质辅助激光解吸/电离(MALDI)质谱成像作为空间分辨分子分析的强大工具出现,提供了克服与合并样品分析相关的主要限制的可能性(Caprioli等,1997; McDonnell and Heeren,2007年)。

损坏左侧调节电路在额颞痴呆中产生更大的积极情绪反应性
积极的情绪是指一个情感家庭,其中包括幸福,娱乐,依恋爱,养育爱,敬畏和热情等(Shiota,Neufeld,Yeung,Yeung,Moser,Moser和Perea,2011年)。这些情绪具有重要的社会功能,促进方法行为,激励社会参与,促进新的社交联系(Fredrickson,2004年),并逆转由负面情绪引起的生理激活(Fredrickson&Levenson,1998)。一定程度的积极情绪反应性被认为是最佳的;太低或太高的水平可能是有问题的。例如,积极情绪过高的基础临床症状,例如阿内迪尼和抑郁症,而过高的水平会导致不适当的人际边界,风险危险和躁狂(Gruber,Harvey,Harvey和Purcell,&Purcell,2011年)。分布在情感上和情绪调节的分布式大脑系统协同行动,以产生观察到的积极情绪反应的水平(通常以面部行为,生理学和主观经验的变化来衡量)。因此,支持积极情绪的神经系统的损伤是否导致情绪柔和或强化的情绪应取决于解剖学损伤的基因座。通常,对情绪产生电路的损害应降低积极的情绪反应性,而对情绪调节电路的损害应削弱抑制作用,从而导致高度带来积极的情绪。长期以来一直在争论积极情绪在大脑中横向的程度。两条证据支持这一结论。While emotion generating sys- tems (i.e., projections from pregenual anterior cingulate cor- tex to the central nucleus of the amygdala, hypothalamus, and brainstem) initiate rapid emotional responses to positive emotional cues ( Saper, 2002 ), emotion regulating systems (i.e., ventrolateral prefrontal cortex, orbitofrontal cortex, dorso- medial prefrontal cortex, and pre/supplementary motor area), with connections to striatum, thalamus, and subthalamic nuclei, promote down-regulation of affective responding in ways that are commensurate with individual goals and the social context ( Aron, 2007; Ochsner & Gross, 2005; Wager, Davidson, Hughes, Lindquist, & Ochsner, 2008 ).有些人认为对积极和负面情绪的感知和表达存在正确的半球优势(Tucker,1981),但其他人则建议左半球在积极情绪中起着主导作用(Davidson&Fox,1982)。先前的研究得出的结论是,左半球损害通常会减少积极的情绪,而右半球损害通常会增加积极的情绪。在WADA的研究中,可以停用右半球(通过单侧氨基脂质注射杏仁钠)但保留左侧的左半球,患者经常表现出乐观和欢笑(Perria,Rosadini和Rossi,&Rossi,&Rossi,1961; Sackeim等,Sackeim等,1982)。同样,许多病变研究,但不是全部(House,Dennis,Warlow,Hawton和Molyneux,1990),发现右半球损伤通常会导致笑声和微笑(Gainotti,1972; Sackeim等,1982)。积极的情绪被认为在右半球损害或功能障碍的范围内持续存在,因为

利用基因组编辑技术生产转基因小鼠的简便方法
细菌免疫。Science。337 : 816-821, 2012。6)Gaj T, Gersbach CA, Barbas CF.: 基于ZFN、TALEN 和CRISPR/Cas 的基因组工程方法。Trends. Biotechnol. 31 : 397-405, 2013。7)Doudna JA, Charpentier E.: 基因组编辑。利用CRISPR-Cas9 进行基因组工程的新前沿。Science。346 : 1258096, 2014。8)Strecker J, Ladha A, Gardner Z 等:利用CRISPR 相关转座酶进行RNA 引导的DNA 插入。Science。 365 :48-53,2019。9)Klompe SE,Vo PLH,Halpin-Healy TS 等:转座子编码的 CRISPR-Cas 系统直接介导 RNA 引导的 DNA 整合。Nature。571 :219-225,2019。10)Jacobi AM,Rettig GR,Turk R 等:用于高效基因组编辑的简化 CRISPR 工具及其向哺乳动物细胞和小鼠受精卵中的精简协议。方法。121-122 :16-28,2017。11)Lino CA,Harper JC,Carney JP 等:CRISPR 的递送:挑战和方法综述。药物递送。 12)Kaneko T.:用于产生和维持有价值动物品系的生殖技术。J. Reprod. Dev. 64:209-215,2018。 13)Mizuno N,Mizutani E,Sato H等:通过腺相关病毒载体通过CRISPR/Cas9介导的基因组编辑实现胚胎内基因盒敲入。iScience。9:286-297,2018。 14)Yoon Y,Wang D,Tai PWL等:利用重组腺相关病毒在小鼠胚胎中精简体外和体内基因组编辑。Nat. Commun. 9 : 412, 2018。15)Takahashi G, Gurumurthy CB, Wada K, 等:GONAD:通过输卵管核酸递送系统进行基因组编辑:一种新型的小鼠微注射独立基因组工程方法。Sci. Rep. 5 : 11406, 2015。16)Sato M, Ohtsuka M, Nakamura S.:输卵管内滴注溶液作为在体内操纵植入前哺乳动物胚胎的有效途径。New Insights into Theriogenology, InTechOpen, London, 2018, pp 135-150。 17)Sato M,Takabayashi S,Akasaka E 等:基因组编辑试剂在小鼠生殖细胞、胚胎和胎儿体内靶向递送的最新进展和未来展望。Cells。9:799,2020。18)Alapati D,Zacharias WJ,Hartman HA 等:宫内基因编辑治疗单基因肺疾病。Sci. Transl. Med。11:eaav8375,2019。19)Nakamura S,Ishihara M,Ando N 等:基因组编辑成分经胎盘递送导致中期妊娠小鼠胎儿胚胎心肌细胞突变。IUBMB life。 20)Sato T, Sakuma T, Yokonishi T 等:利用 TALEN 和双切口 CRISPR/Cas9 在小鼠精原干细胞系中进行基因组编辑。Stem Cell Reports。5:75-82,2015。21)Wu Y, Zhou H, Fan X 等:通过 CRISPR-Cas9 介导的基因编辑纠正小鼠精原干细胞中的一种遗传疾病
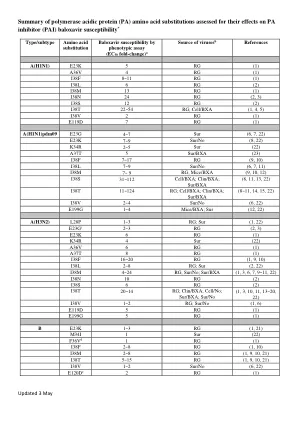
摘要 - 聚合酶 - 酸性 - (PA) - 蛋白质 - 氨基 -
* PA中的其他氨基酸取代,在参考文献1(Omoto S等,2018)和#2(Hashimoto T等,2020年)中研究了Baloxavir易感性没有变化的其他氨基酸取代。通过基于细胞培养的测定法评估(焦点,斑块或屈服分析,高含量成像中和(提示)和ViroDot分析)。EC 50倍变化。b细胞,细胞培养;临床试验;小鼠,鼠标模型; RG,反向遗传学; SUR,监视研究; BXA,在Baloxavir压力下选出的取代;不,Baloxavir不使用。c e23g(T0831)。通过表型测定测试了带有E23G的RG病毒。d对应于A36V A型A型PA中的A36V。 E对应于A型A型PA中的E119D。参考文献1。Omoto S,Speranzini V,Hashimoto T,Noshi T,Yamaguchi H,Kawai M,Kawaguchi K,Uehara T,Shishido T,Naito A,Naito A,Cusack S.2018。通过核酸内切酶抑制剂Baloxavir maroxil诱导的流感病毒变体的表征。SCI REP 8:9633。2。Hashimoto T,Baba K,Inoue K,Okane M,Hata S,Shishido T,Naito A,Wildum S,Omoto S.2020。在Baloxavir Marboxil的临床试验中检测到的流感病毒的三聚体RNA聚合酶复合物中氨基酸取代的全面评估。流感其他呼吸病毒DOI:10.1111/irv.12821。3。ince WL,Smith FB,O'Rear JJ,Thomson M.2020。J Infect DIS 222:957-961。 4。 2018。J Infect DIS 222:957-961。4。2018。治疗 - 伴随流感病毒聚合酶酸性取代率与Balosavir Maroxavir Marboxil试验中的i38中的i38中的酸性取代相关。Noshi T, Kitano M, Taniguchi K, Yamamoto A, Omoto S, Baba K, Hashimoto T, Ishida K, Kushima Y, Hattori K, Kawai M, Yoshida R, Kobayashi M, Yoshinaga T, Sato A, Okamatsu M, Sakoda Y, Kida H, Shishido T, Naito A.Baloxavir酸的体外表征,Baloxavir酸是一种流感病毒聚合酶PA亚基的第一类帽依赖性内切酶抑制剂。抗病毒Res 160:109-117。5。Takashita E,Morita H,Ogawa R,Nakamura K,Fujisaki S,Shirakura M,Kuwahara T,Kishida N,Watanabe S,Odagiri T.2018。流感病毒对新型帽依赖性核酸内切酶抑制剂baloxavir maroxil的敏感性。前微生物9:3026。6。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。 2019。 评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。 欧元监视24:1800666。 7。 Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A. 2020。 全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。 抗病毒Res 175:104718。 8。 2020。Gubareva LV,Mishin VP,Patel MC,Chesnokov A,Nguyen HT,De La Cruz J,Spencer S,Spencer S,Campbell AP,Sinner M,Reid H,Reid H,Garten R,Katz JM,Katz JM,Fry AM,Barnes J,Barnes J,Wentworth DE。2019。评估在2016/17和2017/18季节在美国循环的流感病毒的Baloxavir敏感性。欧元监视24:1800666。7。Takashita E, Daniels RS, Fujisaki S, Gregory V, Gubareva LV, Huang W, Hurt AC, Lackenby A, Nguyen HT, Pereyaslov D, Roe M, Samaan M, Subbarao K, Tse H, Wang D, Yen HL, Zhang W, Meijer A.2020。全球关于人流感病毒对神经氨酸酶抑制剂和cap依赖性核酸内切酶抑制剂Baloxavir的敏感性的更新,2017- 2018年。抗病毒Res 175:104718。8。2020。Takashita E, Abe T, Morita H, Nagata S, Fujisaki S, Miura H, Shirakura M, Kishida N, Nakamura K, Kuwahara T, Mitamura K, Ichikawa M, Yamazaki M, Watanabe S, Hasegawa H, Influenza Virus Surveillance Group of J.流感A(H1N1)PDM09病毒,由于未经Baloxavir治疗的儿童检测到PA E23K替代而表现出对Baloxavir的敏感性降低。抗病毒Res 180:104828。 9。 Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。 2019。 亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。 抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 180:104828。9。Koszalka P,Tilmanis D,Roe M,Vijaykrishna D,Hurt AC。2019。亚太地区流感病毒的Baloxavir Marboxil易感性,2012- 2018年。抗病毒Res 164:91-96。 10。 Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。 2020。 流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。 Proc Natl Acad Sci U S A 117:8593-8601。 11。 Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。 2020。 季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。 J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。抗病毒Res 164:91-96。10。Jones JC,Pascua PNQ,Fabrizio TP,Marathe BM,Seiler P,Barman S,Webby RJ,Webster RG,Govorkova EA。2020。流感和B病毒具有降低的Baloxavir敏感性显示器的体外适应性减弱,但保留了雪貂的可传播性。Proc Natl Acad Sci U S A 117:8593-8601。11。Chesnokov A,Patel MC,Mishin VP,De La Cruz JA,Lollis L,Nguyen HT,Dugan V,Wentworth DE,Gubareva LV。2020。季节性流感A病毒的复制适应性,对Baloxavir的敏感性降低。J Infect DIS 221:367-371。 12。 Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。J Infect DIS 221:367-371。12。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y. 2020。 Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Kiso M,Yamayoshi S,Murakami J,Kawaoka Y.2020。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。 J Infect Dis 221:1699-1702。 13。 Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Baloxavir Marboxil治疗感染了流感病毒的裸小鼠。J Infect Dis 221:1699-1702。13。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。 J Infect DIS 222:121-125。 14。 J Infect DIS 221:63-70。 15。 2020。 16。Sato M,Takashita E,Katayose M,Nemoto K,Sakai N,Hashimoto K,HosoyaM.2020。J Infect DIS 222:121-125。14。J Infect DIS 221:63-70。15。2020。16。在2018-2019流感季节治疗流感A的儿童后,检测Baloxavir Marboxil易感性降低的变体。Checkmahomed L,M'Hamdi Z,Carbonneau J,Venable MC,Baz M,Abed Y,Boivin G.2020。抗性抗性聚合酶酸I38T取代对当代流感A(H1N1)PDM09和A(H3N2)菌株的适应性的影响。Imai M, Yamashita M, Sakai-Tagawa Y, Iwatsuki-Horimoto K, Kiso M, Murakami J, Yasuhara A, Takada K, Ito M, Nakajima N, Takahashi K, Lopes TJS, Dutta J, Khan Z, Kriti D, van Bakel H, Tokita A, Hagiwara H, Izumida N,Kuroki H,Nishino T,Wada N,Koga M,Adachi E,Jubishi D,木谷H,Kawaoka Y.流感A的变体降低了对日本患者分离的Baloxavir敏感性的变体,并通过呼吸道液滴进行拟合。NAT微生物5:27-33。 Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza VirusNAT微生物5:27-33。Takashita E, Kawakami C, Morita H, Ogawa R, Fujisaki S, Shirakura M, Miura H, Nakamura K, Kishida N, Kuwahara T, Mitamura K, Abe T, Ichikawa M, Yamazaki M, Watanabe S, Odagiri T, On Behalf Of The Influenza Virus