XiaoMi-AI文件搜索系统
World File Search Systemaavs

实施CRISPR/CAS9基因组编辑以产生鼠肺癌模型,描绘了人类疾病的突变景观
肺癌是全球最常见的癌症,也是男性和女性癌症相关死亡的主要原因。 尽管发展了新的治疗干预措施,但非小细胞肺癌(NSCLC)患者的5年生存率仍然很低,这表明需要进行新的治疗。 改善转化研究的一种策略是替代模型的发展,反映了肺癌患者鉴定出的体细胞突变,因为这些影响治疗反应。 随着CRISPR介导的基因组编辑的出现,基因删除以及点突变的定位集成,使我们能够比以往任何时候都更详细地对人类恶性肿瘤进行建模。 在这里,我们报告说,通过使用TRP53和KRAS的CRISPR / CAS9介导的靶向,我们概括了经典的Murine NSCLC型号TRP53 plp / plp / pl:lsl-kras g 12 d / wt。 与TRP53频率:LSL -KRAS G 12 d / wt-衍生的肿瘤有关形态,标记表达和转录pro填充物无法区分。 我们证明了CRISPR在体内肿瘤建模的适用性,并改善了使用常规的基因工程小鼠模型的需求。 此外,肿瘤的发作不仅在本构的CAS9表达中,而且在野生型动物中通过感染了肺上皮细胞,其中两个离散的AAV编码了CRISPR机械的不同部分。 虽然传统的鼠标模型需要大量的饲养来整合新的遗传特征,以肺癌是全球最常见的癌症,也是男性和女性癌症相关死亡的主要原因。尽管发展了新的治疗干预措施,但非小细胞肺癌(NSCLC)患者的5年生存率仍然很低,这表明需要进行新的治疗。改善转化研究的一种策略是替代模型的发展,反映了肺癌患者鉴定出的体细胞突变,因为这些影响治疗反应。随着CRISPR介导的基因组编辑的出现,基因删除以及点突变的定位集成,使我们能够比以往任何时候都更详细地对人类恶性肿瘤进行建模。在这里,我们报告说,通过使用TRP53和KRAS的CRISPR / CAS9介导的靶向,我们概括了经典的Murine NSCLC型号TRP53 plp / plp / pl:lsl-kras g 12 d / wt。与TRP53频率:LSL -KRAS G 12 d / wt-衍生的肿瘤有关形态,标记表达和转录pro填充物无法区分。我们证明了CRISPR在体内肿瘤建模的适用性,并改善了使用常规的基因工程小鼠模型的需求。此外,肿瘤的发作不仅在本构的CAS9表达中,而且在野生型动物中通过感染了肺上皮细胞,其中两个离散的AAV编码了CRISPR机械的不同部分。虽然传统的鼠标模型需要大量的饲养来整合新的遗传特征,以

2025; 15(4):1338-1352。 doi:10.7150/thno.100451研究论文RPS23RG1抑制MDGA2的Sort1介导的溶酶体降解以防止自动
基本原理:含有糖基磷脂酰肌醇锚固2(MDGA2)的突触蛋白MAM结构域的突变与自闭症谱系障碍(ASD)有关。因此,阐明MDGA2的调节机制可以帮助开发有效的ASD治疗方法。方法:进行液相色谱串联质谱法以鉴定与RPS23RG1和MDGA2相互作用的蛋白质,然后进行共免疫沉淀测定,以确认蛋白质蛋白质蛋白相互作用。RPS23RG1和Sort1水平被siRNA下调,以研究其对MDGA2降解的影响,并进行了免疫印迹和免疫染色测定的其他应用。溶酶体分离,以进一步确定MDGA2的溶酶体降解。RPS23RG1基因敲除小鼠和MDGA2 +/-小鼠进行各种行为测试,以研究其ASD样表型。在RPS23RG1敲除小鼠中递送表达MDGA2的AAVS,RPS23RG1衍生的肽在MDGA2 +/-小鼠中递送以研究其救援效果。结果:我们发现RPS23RG1和Sort1都与MDGA2相互作用。mdga2主要通过Sort1介导的溶酶体降解途径降解。RPS23RG1与Sort1竞争MDGA2结合以抑制MDGA2降解。此外,我们表明RPS23RG1敲除小鼠表现出降低的MDGA2水平和类似ASD的行为,而MDGA2水平的恢复会减弱RPS23RG1 KO小鼠的社会缺陷。此外,我们确定了用于MDGA2相互作用的RPS23RG1的关键区域,发现源自该区域的肽不仅结合MDGA2并促进MDGA2水平,而且还挽救了MDGA2 +/-小鼠中的社会缺陷。结论:我们的发现突出了RPS23RG1在拮抗MDGA2的Sort1介导的溶酶体降解中的关键作用,并提出了靶向RPS23RG1-MDGA2轴以用MDGA2缺乏处理ASD的潜力。

慢病毒介导的 CRISPR/Cas9 递送可降低小鼠青光眼模型中的眼压
肌动蛋白 ( MYOC ) 突变是已知的原发性开角型青光眼的主要遗传原因,约占所有病例的 4%。MYOC 突变会导致功能获得性表型,其中突变的肌动蛋白会在内质网 (ER) 中积聚,导致 ER 应激和小梁网 (TM) 细胞死亡。因此,在基因组水平上敲除肌动蛋白是永久治愈该疾病的理想策略。我们之前已成功利用 CRISPR/Cas9 基因组编辑通过腺病毒 5 (Ad5) 靶向 MYOC。但是,Ad5 不是适合临床使用的载体。在这里,我们试图确定腺相关病毒 (AAV) 和慢病毒 (LV) 靶向 TM 的功效。首先,我们通过玻璃体内 (IVT) 和前房 (IC) 注射检查了单链 (ss) 和自互补 (sc) AAV 血清型以及表达 GFP 的 LV 的 TM 趋向性。我们观察到 LV_ GFP 表达对通过 IVT 途径注射的 TM 更具特异性。色氨酸突变体 scAAV2 的 IC 注射显示 TM 中 GFP 的显著表达。然而,在睫状体和视网膜中也观察到了强劲的 GFP 表达。我们接下来构建了表达 Cas9 和靶向 MYOC (crMYOC) 的引导 RNA (gRNA) 的慢病毒颗粒,并用 LV_cr MYOC 转导稳定表达突变型肌动蛋白的 TM 细胞可显著减少肌动蛋白积累及其相关的慢性 ER 应激。在 Tg-MYOC Y437H 小鼠中单次 IVT 注射 LV_cr MYOC 可减少 TM 中的肌动蛋白积累并显著降低升高的眼压。总之,我们的数据表明,LV_cr MYOC 靶向 TM 中的 MYOC 基因编辑并挽救了肌动蛋白相关青光眼的小鼠模型。

与腺相关的载体分配的CRISPR/SACAS9系统减少了猫的白血病病毒在体外生产
摘要:猫白血病病毒(FELV)是全球猫的逆转录病毒。高病毒载量与进行性感染和宿主死亡有关,这是由于FELV相关疾病而导致的。相比之下,在感染回归的猫中,可以观察到低病毒负荷,有效的免疫反应和更好的临床结局。我们假设通过使用CRISPR/ SA CAS9辅助基因治疗降低逐渐感染的猫的病毒载荷,可以允许该猫的免疫系统将感染引导到回归结果。在朝着这一目标的一步中,本研究评估了不同的腺相关载体(AAV),以使其能够将基因编辑系统传递到猫科动物细胞中,然后研究针对FELV FORIRUS内不同站点的CRISPR/ SA CAS9。九种天然AAV血清型,两种AAV杂种菌株和ANC80L65(硅中的ANC80L65预测AAV祖先)的测试是针对感染不同猫线细胞系和猫科动物原代细胞的潜力。AAV-DJ揭示了较高的感染效率,因此在随后的转导实验中使用。使用CRISPR/ SA CAS9系统的引入12个选定的FELV Profirus站点,由T7核酸内切酶1(T7E1)确认,并通过分解(TIDE)分析来跟踪Indels。在高度保守的GAG和POL区域中发现了非同源末端加入(NHEJ)的最高百分比(最高80%)。随后的转导实验,使用AAV-DJ确认了indel的形成,并显示了某些靶标的FELV P27抗原的显着降低。在体外使用CRISPR/ SA CAS9方法时,FELV病毒的靶向是有效的。观察到的靶向病毒靶向的程度是否足以提供逐渐感染的FELV感染的猫来克服感染的手段,需要在体内进一步研究。

基因组编辑器的体内靶向非病毒递送
RNA 引导的 CRISPR-Cas 酶因其功效、灵活性和易用性而被广泛用于基因组编辑 [已在其他地方进行综述 (1, 2)]。虽然 Cas9 等 CRISPR 蛋白已经在临床试验中显示出良好的前景,但对人类基因组造成永久性改变的现实意味着安全性至关重要。在基因组层面,Cas9 的特异性已通过预测脱靶位点的方法 (3, 4) 和分子工程来产生高保真度蛋白质 (5) 进行了优化。然而,一项将提高基因组编辑的实用性和安全性的关键发展是能够将 CRISPR-Cas 基因组编辑机制专门递送到患者体内所需的细胞类型、组织或器官。对于许多遗传疾病,只有一小部分细胞或特定器官表现出疾病的表型迹象,因此将成为基因组编辑的预期目标。对非预期细胞或器官进行基因组编辑可能会增加意外治疗结果的风险,此外还会因更高的剂量要求而增加制造成本。目前,CRISPR-Cas 基因组编辑器的靶向递送仍然是成功实现基因组编辑临床转化的重要未满足需求。病毒载体缺乏其天然基因组和复制能力,是基因治疗和最近的 CRISPR-Cas 基因组编辑的一种有吸引力的递送策略[在其他地方进行了综述 (6)]。最广泛使用的病毒载体是逆转录病毒和腺相关病毒 (AAV) (7, 8)。慢病毒载体是逆转录病毒的一个亚型,在基因组整合后表达较大的转基因 (~ 10 kb),而 AAV 表达较小的转基因 (~ 4.7 kb),来自长寿命的附加体;这两种病毒载体都能够转导分裂细胞和非分裂细胞。假型慢病毒载体 (9)、新 AAV 趋向性工程 (10) 和组织特异性启动子使用的进展使得这些技术能够实现细胞特异性递送。然而,病毒递送也引入了

基因组编辑器的体内靶向非病毒递送
RNA 引导的 CRISPR-Cas 酶因其功效、灵活性和易用性而被广泛用于基因组编辑 [已在其他地方进行综述 (1, 2)]。虽然 Cas9 等 CRISPR 蛋白已经在临床试验中显示出良好的前景,但对人类基因组造成永久性改变的现实意味着安全性至关重要。在基因组层面,Cas9 的特异性已通过预测脱靶位点的方法 (3, 4) 和分子工程来产生高保真度蛋白质 (5) 进行了优化。然而,一项将提高基因组编辑的实用性和安全性的关键发展是能够将 CRISPR-Cas 基因组编辑机制专门递送到患者体内所需的细胞类型、组织或器官。对于许多遗传疾病,只有一小部分细胞或特定器官表现出疾病的表型迹象,因此将成为基因组编辑的预期目标。对非预期细胞或器官进行基因组编辑可能会增加意外治疗结果的风险,此外还会因更高的剂量要求而增加制造成本。目前,CRISPR-Cas 基因组编辑器的靶向递送仍然是成功实现基因组编辑临床转化的重要未满足需求。病毒载体缺乏其天然基因组和复制能力,是基因治疗和最近的 CRISPR-Cas 基因组编辑的一种有吸引力的递送策略[在其他地方进行了综述 (6)]。最广泛使用的病毒载体是逆转录病毒和腺相关病毒 (AAV) (7, 8)。慢病毒载体是逆转录病毒的一个亚型,在基因组整合后表达较大的转基因 (~ 10 kb),而 AAV 表达较小的转基因 (~ 4.7 kb),来自长寿命的附加体;这两种病毒载体都能够转导分裂细胞和非分裂细胞。假型慢病毒载体 (9)、新 AAV 趋向性工程 (10) 和组织特异性启动子使用的进展使得这些技术能够实现细胞特异性递送。然而,病毒递送也引入了
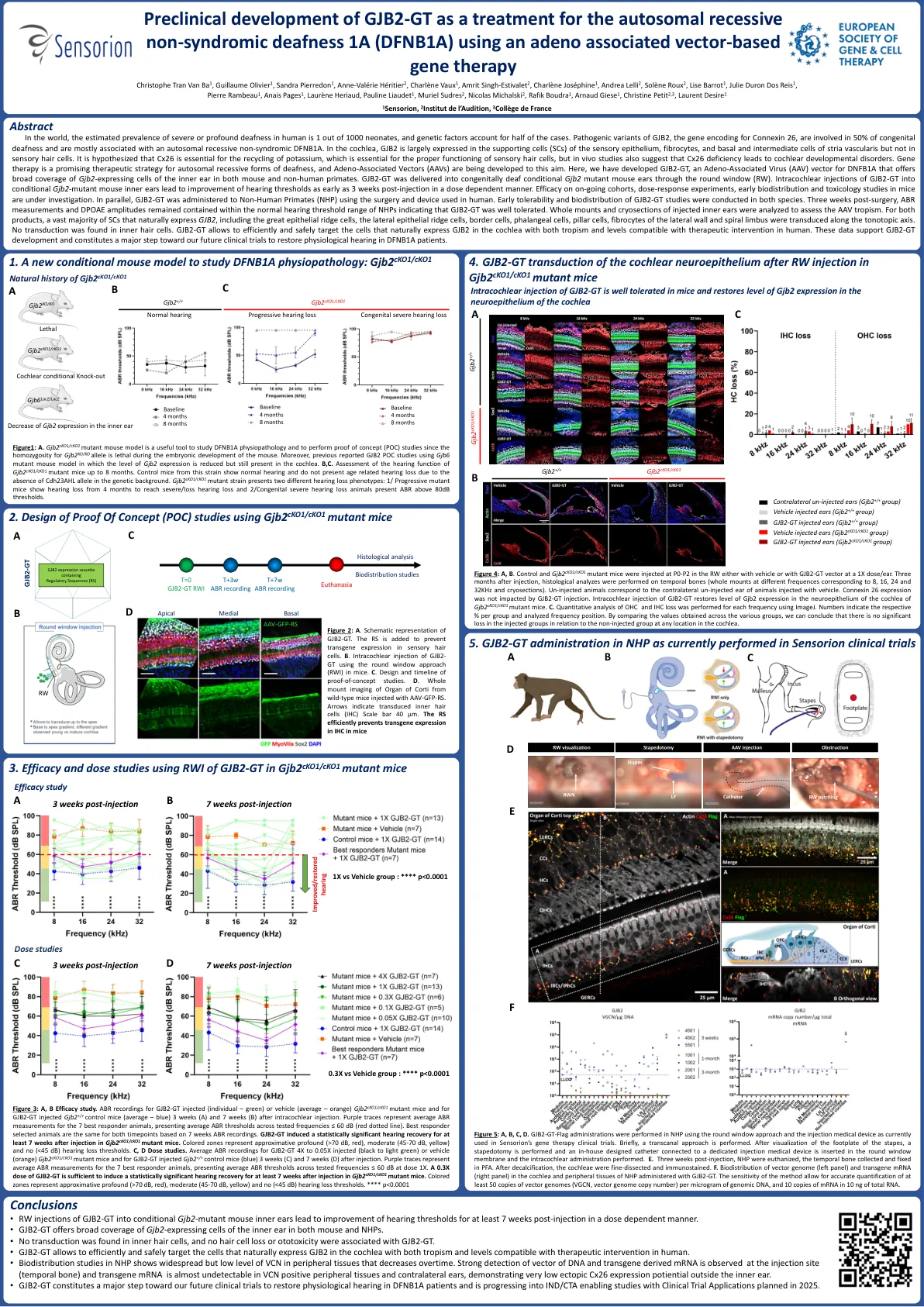
GJB2-GT作为... 的临床前开发 sens-501常染色体隐性非... 的基因治疗
在世界上,人类严重或深刻的耳聋的估计患病率是1000名新生儿中的1个,遗传因素占了一半的病例。 GJB2的致病变异,编码连接蛋白26的基因,涉及50%的先天性耳聋,主要与常染色体隐性遗传性非伴有伴有伴有dfnb1a有关。 在耳蜗中,GJB2在感官上皮,纤维细胞,基底和中间细胞的血管毛血管的辅助细胞(SC)中主要表达,但在感觉毛细胞中却没有。 据推测,CX26对于钾的回收至关重要,这对于感觉毛细胞的正确功能至关重要,但是体内研究还表明CX26缺乏会导致耳蜗发育障碍。 基因疗法是一种有前途的聋哑形式的有前途的治疗策略,并且正在为此目的而开发与腺相关的载体(AAV)(AAVS)。 在这里,我们开发了GJB2-GT,这是DNFB1A的腺相关病毒(AAV)载体(AAV)载体,可在小鼠和非人类灵长类动物中均提供GJB2表达内耳gjb2表达细胞的广泛覆盖范围。 gjb2-gt通过圆形窗口(RW)传递到先天性聋哑的GJB2突变小鼠耳朵中。 对条件GJB2的gjb2-GT对有条件的小鼠内耳的注射会导致听力阈值在注射后3周以剂量依赖的方式改善。 对持续的队列,剂量反应实验,早期生物分布和毒理学研究的功效正在研究中。 并行,使用人类使用的手术和装置将GJB2-GT用于非人类灵长类动物(NHP)。在世界上,人类严重或深刻的耳聋的估计患病率是1000名新生儿中的1个,遗传因素占了一半的病例。GJB2的致病变异,编码连接蛋白26的基因,涉及50%的先天性耳聋,主要与常染色体隐性遗传性非伴有伴有伴有dfnb1a有关。在耳蜗中,GJB2在感官上皮,纤维细胞,基底和中间细胞的血管毛血管的辅助细胞(SC)中主要表达,但在感觉毛细胞中却没有。据推测,CX26对于钾的回收至关重要,这对于感觉毛细胞的正确功能至关重要,但是体内研究还表明CX26缺乏会导致耳蜗发育障碍。基因疗法是一种有前途的聋哑形式的有前途的治疗策略,并且正在为此目的而开发与腺相关的载体(AAV)(AAVS)。在这里,我们开发了GJB2-GT,这是DNFB1A的腺相关病毒(AAV)载体(AAV)载体,可在小鼠和非人类灵长类动物中均提供GJB2表达内耳gjb2表达细胞的广泛覆盖范围。gjb2-gt通过圆形窗口(RW)传递到先天性聋哑的GJB2突变小鼠耳朵中。对条件GJB2的gjb2-GT对有条件的小鼠内耳的注射会导致听力阈值在注射后3周以剂量依赖的方式改善。对持续的队列,剂量反应实验,早期生物分布和毒理学研究的功效正在研究中。并行,使用人类使用的手术和装置将GJB2-GT用于非人类灵长类动物(NHP)。在这两种物种中均进行了GJB2-GT研究的早期耐受性和生物分布。 手术后三周,ABR测量和DPOAE振幅保留在NHP的正常听力阈值范围内,表明GJB2-GT耐受性良好。 分析了注射的内耳的整个安装和冷冻切片,以评估AAV的偏向主义。 对于这两种产品,绝大多数自然表达GJB2的SC,包括大上皮脊细胞,侧皮脊细胞,边界细胞,圆锥细胞,柱状细胞,侧壁的纤维细胞,侧壁和螺旋状肢体的纤维细胞沿着负轴轴线进行传播。 在内毛细胞中未发现转导。 GJB2-GT允许有效,安全地靶向自然表达GJB2在耳蜗中的细胞,并具有与人类治疗干预兼容的水平。 这些数据支持GJB2-GT开发,并构成了我们未来的临床试验迈出的重大步骤,以恢复DFNB1A患者的生理听力。在这两种物种中均进行了GJB2-GT研究的早期耐受性和生物分布。手术后三周,ABR测量和DPOAE振幅保留在NHP的正常听力阈值范围内,表明GJB2-GT耐受性良好。分析了注射的内耳的整个安装和冷冻切片,以评估AAV的偏向主义。对于这两种产品,绝大多数自然表达GJB2的SC,包括大上皮脊细胞,侧皮脊细胞,边界细胞,圆锥细胞,柱状细胞,侧壁的纤维细胞,侧壁和螺旋状肢体的纤维细胞沿着负轴轴线进行传播。在内毛细胞中未发现转导。GJB2-GT允许有效,安全地靶向自然表达GJB2在耳蜗中的细胞,并具有与人类治疗干预兼容的水平。这些数据支持GJB2-GT开发,并构成了我们未来的临床试验迈出的重大步骤,以恢复DFNB1A患者的生理听力。
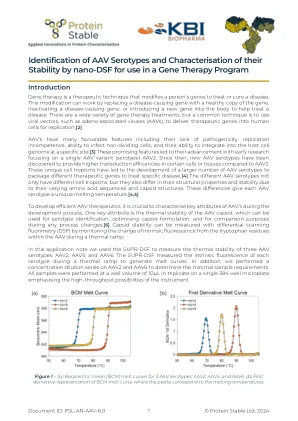
识别AAV-Serotypes and-thacteration-of-...
基因治疗是一种治疗技术,可修饰一个人的基因治疗或治愈疾病。这种修饰可以通过用健康的基因副本代替引起疾病的基因,使疾病的基因失活或将新基因引入体内以帮助治疗疾病。有多种基因疗法治疗,但是一种常见的技术是使用病毒载体,例如腺相关病毒(AAV),将治疗基因传递到人类细胞中进行复制。[2] AAV具有许多有利的特征,包括缺乏致病性,复制无能力,感染非分散细胞的能力以及在特定部位中整合到宿主细胞基因组中的能力。[3]这些有前途的特征导致了它们的进步,早期研究重点是单个AAV变体(血清型)AAV2。从那时起,与AAV2相比,已经发现了新的AAV血清型在某些细胞或组织中提供较高的转导效率。这些独特的细胞偏向主义已导致大量AAV血清型的发展来包装不同的治疗基因来治疗特定疾病。[4]不同的AAV血清型不仅具有不同的细胞向量,而且由于其不同的氨基酸序列和衣壳结构,它们的结构特性和稳定性也有所不同。这些差异使每种AAV血清型都具有独特的熔化温度。[4,5]为了开发有效的AAV疗法,在开发过程中表征AAV的关键属性至关重要。SUPR-DSF在热坡道期间测量了每种血清型的固有荧光,以产生熔体曲线。一个关键属性是AAV CAPSID的热稳定性,可用于血清型识别,优化衣壳配方以及在任何过程更改中进行比较。[6]可以通过在热坡道期间从AAV中的色氨酸残基中监测固有荧光的变化,可以通过差异扫描荧光法(DSF)测量衣壳稳定性。在此应用程序注释中,我们使用SUPR-DSF来测量三种AAV血清型的热稳定性:AAV2,AAV5和AAV6。此外,我们在AAV2和AAV6上进行了浓度稀释系列,以确定最小样品要求。所有样品均以10µl的井体积为10µl,在单个384孔微孔板上一式三份,强调该仪器的高通量可能性。

ASHG 2023 全体会议摘要
使用 CRISPR/Cas9 进行神经遗传疾病的基因编辑面临难以穿过血脑屏障、渗透性有限和治疗窗口狭窄的挑战。虽然改良的腺相关病毒 (AAV) 克服了其中一些障碍,但由于 Cas9 蛋白的长期存在,它们的免疫原性和更高的脱靶效应风险限制了它们对人类神经遗传疾病的转化价值。为了解决这个问题,我们开发了一种创新的非病毒递送工具,使用与 Cas9 蛋白和 sgRNA 结合的化学修饰核糖核蛋白 (RNP) (cRNP-Cas9/sgRNA,cRNPcg)。由于其尺寸小 (12um),cRNPcg 能够有效渗透到大脑中的神经元细胞中,而瞬时 Cas9 蛋白大大降低了脱靶效应的风险。我们在体外和体内测试了 cRNPcg 对 Angelman 综合征 (AS) 的疗效,这是一种由神经元和母体特异性 UBE3A 基因表达缺陷引起的神经发育障碍。父系染色体中 UBE3A 的抑制表达由父系表达的非编码 UBE3A 反义转录本 (UBE3A-ATS) 介导。通过反义寡核苷酸 (ASO) 灭活 UBE3A-ATS 在正在进行的 1/2 期临床试验中显示出积极的临床效果。然而,ASO 的短暂作用需要每月鞘内注射,这对作为标准临床治疗方法提出了挑战。我们设计的 cRNPcg 系统可选择性地灭活 Ube3a-ATS 表达,并可能通过单次治疗实现永久性治疗效果。使用 Ube3a-YFP 报告小鼠,我们观察到高基因编辑效率(>75% 靶向细胞)和广泛的脑渗透。我们给新生儿 (P1-2) 和 P21 AS Ube3 a m-/p+ 模型鞘内注射了 cRNPcg,观察到 Ube3a-ATS 显著降低,并且 Ube3a 重新激活至正常水平的 30%,遍及皮质、海马和小脑。因此,这种治疗显著改善了多个行为领域,包括运动功能、焦虑样行为、学习和记忆,并且还延长了成年 AS Ube3 a m-/p+ 小鼠化学诱发的肌阵挛和强直性癫痫发作的潜伏期。重要的是,我们没有观察到与 cRNPcg 相关的任何急性或慢性毒性。此外,我们发现 cRNPcg 有效地重新激活了 AS 患者 hIPSC 衍生的神经祖细胞中父系染色体上的 UBE3A 表达,这些神经祖细胞存在 15q11-q13 的大量母系缺失。总之,我们的结果表明,cRNPcg 是一个将 CRISPR/Cas9 基因编辑传递到大脑的创新平台,具有广泛的应用和治疗许多其他神经遗传疾病的潜力。