XiaoMi-AI文件搜索系统
World File Search Systemactivation

支持信息 DNA 损伤检查点激活...
引物名称 序列 oEAM20 cttagtcgactaaggaggtatacatatgagtaaagaatctattatttttgtcg oPEB857 gaattagcttgcatgcggctagcttactatcttacagttgctaattcatatg oPEB3F gctagccgcatgcaagctaattcg oPEB259 atgtatacctccttagtcgactaagcttaattgttatccgctcacaattacacacattatgccacacc ttgtagata oPEB866 tttatgcagcaatggcaagaac oPEB867 gccgactcaaacatcaaatcttac oEAM21 gaattagcttgcatgcggctagcttattcatatgcatcctgatgcttctttttc oEAM22 gaattagcttgcatgcggctagcttaatgcttctttttcaatgggatcactaac oEAM23 gaattagcttgcatgcggctagcttatgggatcactaactcatcaccc oEAM28 cttagtcgactaaggaggtatacatatgcggcagctgtttgctttaattg oEAM29 gaattagcttgcatgcggctagcttattacagataaacagttttttgagctgtttc oEAM42 tcccggtgaaggtcaaggacaaggccaaatgaacccgggtcaagaccgaga oEAM43 gttgtaaaacgacggccgaattcttagttatcaattttcatcttcctttttagcagct oEAM44 tcccggtgaaggtcaaggacaaggccaaatggagtttgtcattggattaattg oEAM45 gttgtaaaacgacggccgaattcttagttaactaagcggatatgtcagctttgat oEAM46 tcccggtgaaggtcaaggacaaggccaaatgagtatgaaaaataaactgaaaaactt oEAM47 gttgtaaaacgacggccgaattcttagttattaaccacctctgatgttcgtcttc oEAM48 tcccggtgaaggtcaaggacaaggccaattgaattttccagggaacgaacg oEAM49 gttgtaaaacgacggccgaattcttagttaactacttgctcttcttctccacatt oEAM50 tcccggtgaaggtcaaggacaaggccaaatgagcaatttagcttaccaacca oEAM51 gttgtaaaacgacggccgaattcttagttatcattcctgtatgtttttcacttttttatc oEAM52 tcccggtgaaggtcaaggacaaggccaaatgcggcagctgtttgctttaatt oEAM53 gttgtaaaacgacggccgaattcttagttattacagataaacagtttttttgagctg oEAM54 tcccggtgaaggtcaaggacaaggccaaatgccattaacgccaaatgatattc oEAM55 gttgtaaaacgacggccgaattcttagttattattccttttcctcaaatacagcg oEAM56 tcccggtgaaggtcaaggacaaggccaaatgccaaaaaaagaataaatttatgaatagag oEAM57 gttgtaaaacgacggccgaattcttagttattaatcaggatttttaaacttaaccttg oPEB1021 attatgccgcatctgtccaact oPEB1022 gcaaggcgattaagttgggtaa oPEB1024 ttctcgccggatgtactggaaac oPEB1025 tggcttaactatgcggcatcaga oPEB1034 ctagagggtgaaggtcaaggacaaggccaaatgagtaaagaatctattatttttgtcggt oPEB1035 gtactgagagtgcaccatattacttagttaactatcttacagttgctaattcatatgca oPEB1041 atcccggtgaaggtcaaggacaaggccaattgaaacggcaattaaaactgttttttattg oPEB1042 gttgtaaaacgacggccgaattcttagttattacatttctttttcagtgtttcaattgc oPEB1058 tcccggtgaaggtcaaggacaaggccaaatgagtaaagaatctattattttgtcggt oPEB1059 gttgtaaaacgacggccgaattcttagttaactatctttacagttgctaattcatatgca oPEB1070 tctagagggtgaaggtcaaggacaaggccaaatgaacccgggtcaagaccgaga oPEB1071 actgagagtgcaccatattacttagttatcaattttcatcttcctttttagcagct oPEB1072 tagagggtgaaggtcaaggacaaggccaaatggagtttgtcattggattattaattg oPEB1073 ttgtactgagagtgcaccatattacttagttactaagcggatatgtcagctttgat oPEB1074 tagagggtgaaggtcaaggacaaggccaaatgagtatgaaaaataaactgaaaaactt

KIF4A 激活凝聚素 I 的分子机制
在有丝分裂过程中,凝缩蛋白 I 和 II 复合物将染色质压缩成染色体。染色质驱动蛋白 KIF4A 的缺失会导致凝缩蛋白 I 与染色体的结合减少,但这种表型背后的分子机制尚不清楚。在本研究中,我们发现 KIF4A 通过位于其 C 末端尾部的保守无序短线性基序 (SLiM) 直接与人类凝缩蛋白 I HAWK 亚基 NCAPG 结合。 KIF4A 与 NCAPH N 端和 NCAPD2 C 端的 SLiM 竞争 NCAPG 与重叠位点的结合,后者介导凝聚素 I 中的两种自抑制相互作用。KIF4A SLiM 肽本身就足以刺激凝聚素 I 的 ATPase 和 DNA 环挤压活性。我们在已知的酵母凝聚素相互作用蛋白 Sgo1 和 Lrs4 中发现了类似的 SLiM,它们与酵母凝聚素亚基 Ycg1(与 NCAPG 相当的 HAWK)结合。我们的研究结果以及之前对凝聚素 II 和黏连素的研究证明,SLiM 与 NCAPG 相当的 HAWK 亚基结合是 SMC 复合物中保守的调节机制。

美国心脏月激活指南2023
戴上红色,至少有一个您优先考虑的健康习惯来成为节拍 - 它可能围绕睡眠,压力,体育锻炼,营养或最适合您的任何东西 - 并鼓励其他人也这样做。我们可以帮助我们认识的妇女,并通过养成我们知道工作的健康习惯来减少他们患心血管疾病的风险。

IL-17RD 慢病毒激活颗粒 (m):sc-431369-LAC
成簇的规律间隔短回文重复序列 (CRISPR) 和 CRISPR 相关蛋白 (Cas9) 系统是一种适应性免疫反应防御机制,古细菌和细菌利用该机制来降解外来遗传物质。该机制可以重新用于其他功能,包括哺乳动物系统的基因组工程,例如基因敲除 (KO) (1,2) 和基因激活 (3-5)。CRISPR 激活质粒产品利用与 VP64 激活域融合的 D10A 和 N863A 失活 Cas9 (dCas9) 核酸酶与 sgRNA (MS2) 结合,从而实现特定基因的识别和上调,sgRNA (MS2) 是一种靶向特异性 sgRNA,经过设计可结合 MS2-P65-HSF1 融合蛋白 (5)。这种协同激活介质 (SAM) 转录激活系统* 提供了一个强大的系统,可最大限度地激活内源性基因表达 (5)。

非小细胞肺癌中 BRAF 激活的演变
非小细胞肺癌(NSCLC)是最常见的肺癌亚型,其中约4%具有BRAF激活,可选择靶向治疗。BRAF激活包括V600和非V600突变,融合,重排,框内缺失,插入和共突变。此外,BRAF原发性激活和继发性激活具有不同的生物学表型,医学意义和后续治疗。BRAF原发性激活作为NSCLC的驱动基因在增殖和转移中起着关键作用,而继发性激活介导对其他靶向治疗的获得性耐药,尤其是表皮生长因子酪氨酸激酶抑制剂(EGFR-TKI)。针对不同BRAF激活的治疗选择多种多样。靶向治疗,尤其是双药联合治疗,是一种重要的选择。此外,免疫检查点抑制剂(ICI)将是另一种选择,因为BRAF激活将是ICI治疗的肿瘤反应的阳性生物标志物。目前尚无高级别证据支持将靶向治疗或免疫治疗作为优先推荐。靶向治疗后,BRAF 的进化包括激活 BRAF 的上游、下游和旁路通路。本文将讨论 BRAF 的治疗方式和治疗后进化通路,并提出未来的研究方向。

CRISPR 介导的 αV 整合素亚型激活... - ArTS
神经元分化是一个复杂的过程,其功能障碍会导致脑部疾病。开发新工具以针对神经元分化过程中的特定步骤至关重要,这有利于更好地理解所涉及的分子机制,并最终为神经发育障碍制定有效的治疗策略。通过与细胞外基质蛋白的相互作用,整合素家族的细胞粘附分子通过调节细胞迁移、神经突生长、树突棘形成和突触可塑性,在功能性神经元回路的形成中发挥重要作用。然而,不同的整合素受体如何促进神经元分化的连续阶段仍有待阐明。在这里,我们实施了一个 CRISPR 激活系统来增强特定整合素亚基在神经元分化的体外模型(鼠神经母细胞瘤 Neuro2a 细胞系)中的内源性表达。通过将 CRISPR 激活与形态学和 RT-qPCR 分析相结合,我们表明 α V 家族的整合素是神经元分化的强大诱导剂。此外,我们确定了 α V 整合素在控制神经突生长方面具有亚型特异性作用。虽然 α V β 3 整合素在增殖条件下启动 Neuro2a 细胞的神经元分化,但 α V β 5 整合素似乎负责促进已分化细胞中的复杂树突分化。有趣的是,原代神经元在发育过程中表现出 β 3 和 β 5 整合素亚基的互补表达模式。我们的研究结果揭示了分化过程中 α V 整合素亚型之间存在发育转换,并表明 CRISPRa 及时控制调节 α V 整合素的表达提供了一种促进神经元分化的方法。
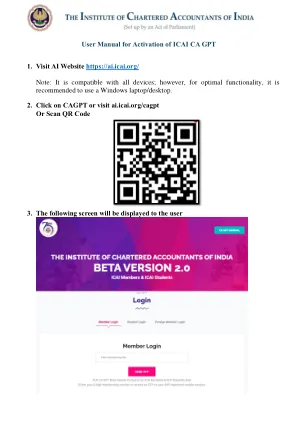
激活ICAI CA GPT
免责声明:ICAI CA GPT是Beta版本,其中所有由ICAI创建和配置的GPT由开放AI平台供电。Beta版本在训练阶段和机器学习中,根据收到的反馈和建议,正在改善及时响应。从ICAI CA GPT内容收到的响应仅用于信息和教育目的,不应被视为合法,会计或税收建议,或者是获得特定于您的企业的建议的替代品。此Beta版本使用知识存储库和培训模型。虽然努力确保准确性,但可能会出现错误。在ICAI CA GPT上的所有内容都以“可用”为基础提供给您的所有内容,无论是否有明示或暗示的保证,包括但不限于任何隐含的适销性,适用于特定目的,准确性和非侵权的ICAI的适合性,对通过此平台的准确性,完整性,货币,货币或可靠性都没有保证。成员使用自己的回复。


小胶质细胞,星形胶质细胞和神经元中的细胞类型特异性激活
相应的作者Dietmar R.神经病理学的Thal实验室O&N IV Hestraat 49 -Bus 1032 3000 Leuven Tel。:0032/16/3-44047电子邮件地址:dietmar.thal@kuleuven.be

量子开关信息能力激活的热力学激活
我们解决了一个新的环境,其中第二定律受到质疑:因果订单的量子叠加中的热量,由所谓的量子开关制定。这种叠加已被证明与通道的通信能力的增加有关,从而显然违反了数据处理不平等,并且有可能将热与寒冷分开。我们分析了此信息能力增加过程的热力学。我们展示了信息能力增加与热力学的兼容。我们表明,如果连续热力学的连续热量遵守热力学的第一和第二定律,则可能确实会增加信息能力,如果将它们放置在不确定的顺序上,此外,只有显着限制的增加才有可能。增加是以消耗热力学资源的代价,即与开关相关的连贯性的自由能。