XiaoMi-AI文件搜索系统
World File Search Systemblot

定量分析和优化哺乳动物细胞中位点特异性蛋白生物结合
摘要:尽管有多种共价蛋白质修饰,但很少有用于定量细胞中蛋白质生物结合的技术。在这里,我们描述了一种通过与Halotag共价键形成在纤维素蛋白生物偶联中量化的新方法。这种方法利用不自然的氨基酸(UAA)诱变选择性地在蛋白质表面上安装小而生物串管的反应性手柄。我们利用了反电子二极管的快速动力学和高选择性 - 评估四嗪苯丙氨酸(TETF)与紧张的反甲环烯 - 氯酸酯(STCO-CA)(STCO-CA)和跨循环链烯(TETR-caclecten)(TETR-CATRE)的反应(TETF)与TETRECANE(TETRE)(TETER-CARORE(TETRE)。生物缀合后,叶绿素配体暴露于释放酶标记,以通过简单的蛋白质印迹分析直接定量生物缀合。我们证明了该工具的多功能性,以快速,准确地确定不同UAA/氯烷烃对的生物缀合效率以及对不同蛋白质的不同位点(包括EGFP和雌激素相关的受体ERR)的不同位点。■简介

kifc1:在RB阳性三阴性乳腺癌患者中使用阿霉素治疗的可靠预后生物标志物与Abemaciclib结合使用
摘要。背景/目标:非洲裔美国人(AA)患者的三阴性乳腺癌(TNBC)患病率和复发风险最大。先前已显示了RB阳性TNBC细胞(MDA-MB-231)中的阿霉素(Dox)和Abemaciclib(Abe)协同作用,以及RB阴性TNBC细胞(MDA-MB-468)中的拮抗作用。在这里,我们评估了类似驱动蛋白样蛋白1(KIFC1)作为TNBC中RB状态的DOX+ABE组合的种族特异性预后生物标志物。材料和方法:在AA种群中搜索TNBC预后生物标志物的文献。MDA-MB-231和MDA-MB-468细胞在72小时内暴露于四个治疗组:1)对照(不含药物的培养基),2)DOX在MDA-MB-231(0.565μm)中的50%抑制浓度下,单独使用MDA-MB-231(0.565μm)和MDA-MB-4-468(0.121μm),和468(0.121μm),和2μmMM(2μm)每个单元线中的相应浓度。KIFC1蛋白表达和时间变化在MDA-MB-231细胞中使用Western blot进行了定量。结果:KIFC1,Kaiso和Annexin A2是文献 - 鉴定出AA特异性TNBC预后生物标志物。kifc1与其他提出的生物标志物无关,

补充信息
补充图 S5。olslc38a4 (SAT) 的消除对青鳉幼虫表型、上皮 Na + 通量和蛋白质表达的影响。 (A),青鳉 Sat 与商用抗 SLC38A4 抗体免疫原肽(针对人类 SLC38A4 的合成肽;序列同源性:74%;ab58785;Abcam Cambridge,英国)的推断序列比对。Western blot 分析 SAT 消除对 Sat 变体蛋白质表达的影响。分别应用了 6 dpf 青鳉幼虫匀浆(从 FW 中的野生型、20‰ SW 中的野生型和 20‰ SW 中的 Sat 变体中收集),并表明商用抗 SLC38A4 抗体可以检测到来自不同青鳉幼虫样本的蛋白质,预期分子量大小约为 56 KDa。 (B) 野生型 (Wt) 和 1 ng SAT MO 注射青鳉胚胎在 20‰ SW 条件下的光学显微镜图像。 (C) 淡水 (FW) 环境下,与野生型和假对照青鳉幼体相比,SAT MO 注射对 6 dpf 青鳉幼体 Na + 通量的影响。值以平均值 ± SD 表示,并使用 Student's t 检验进行比较。当 p < 0.05 时,认为存在显著差异。

NH-硫胺 - UCL发现 - 伦敦大学学院
图2 NHS对ATP动力学的影响。 (a)NHS诱导1(代表n = 6)的二聚化。 (b)暴露于NHS(1μm)viatmrm(20 nm)荧光的SH-SY5Y细胞中的Δψm评估。 (c)条形图量化线索 - 膜电位(Δψm)。 数据显示为平均值±SEM(n = 14)。 * p <0.05,如所示。 (d - e)由Liuminometer记录的代表性痕迹在用线粒体靶向(MIT)和凝结核酸(Cyt)荧光素酶转染的SH-SY5Y细胞中,并用荧光素(100μm)灌注。 在高原上,将用NHS(1μm)挑战细胞,并监测动力学(n = 9)。 (F - G)SH-SY5Y细胞被PGIPZ GFP标记的载体稳定转染(如第2节所述),如果通过(F)中的Western blot分析确认了1个下调。 (g)条显示了1个表达的变化,将1个表达归一化为β-肌动蛋白水平,并表示为平均值±SEM(n = 9)。 * p <0.05,如所示。 (H)响应NACN和IAA处理的MGG荧光变化的代表性痕迹。 (i)条显示了在NaCN(1 mM)和IAA(2 mM)存在下,用NHS1μm处理18-H处理后对应于ATP耗竭的MGG荧光的变化。 数据归一化为未处理的细胞,并表示为平均值±SEM(n = 11)。 * p <0.05,如所示。 * P <0.05,如所示明显不同图2 NHS对ATP动力学的影响。(a)NHS诱导1(代表n = 6)的二聚化。(b)暴露于NHS(1μm)viatmrm(20 nm)荧光的SH-SY5Y细胞中的Δψm评估。(c)条形图量化线索 - 膜电位(Δψm)。数据显示为平均值±SEM(n = 14)。* p <0.05,如所示。(d - e)由Liuminometer记录的代表性痕迹在用线粒体靶向(MIT)和凝结核酸(Cyt)荧光素酶转染的SH-SY5Y细胞中,并用荧光素(100μm)灌注。在高原上,将用NHS(1μm)挑战细胞,并监测动力学(n = 9)。(F - G)SH-SY5Y细胞被PGIPZ GFP标记的载体稳定转染(如第2节所述),如果通过(F)中的Western blot分析确认了1个下调。(g)条显示了1个表达的变化,将1个表达归一化为β-肌动蛋白水平,并表示为平均值±SEM(n = 9)。* p <0.05,如所示。(H)响应NACN和IAA处理的MGG荧光变化的代表性痕迹。(i)条显示了在NaCN(1 mM)和IAA(2 mM)存在下,用NHS1μm处理18-H处理后对应于ATP耗竭的MGG荧光的变化。数据归一化为未处理的细胞,并表示为平均值±SEM(n = 11)。* p <0.05,如所示。* P <0.05,如所示(j和k)然后,用NHS1μM处理后,根据(J)NaCn或(K)IAA评估MGG荧光的增加。

水稻热休克蛋白60-3B通过淀粉颗粒生物合成维持高温下的雄性生育能力
高温对水稻 (Oryza sativa) 的雄性育性有有害影响,但水稻雄配子体免受高温胁迫的机制尚不清楚。在这里,我们分离并鉴定了一种热敏感的雄性不育水稻突变体——热休克蛋白 60-3b (oshsp60-3b),它在最适温度下表现出正常的育性,但随着温度升高育性降低。高温会干扰 oshsp60-3b 花药中花粉淀粉颗粒的形成和活性氧 (ROS) 清除,导致细胞死亡和花粉败育。与突变体表型一致,OsHSP60-3B 在热休克反应中迅速上调,其蛋白质产物定位于质体。至关重要的是,OsHSP60-3B 的过表达增强了转基因植物花粉的耐热性。我们证实 OsHSP60-3B 与质体中的粉质胚乳 6 (FLO6) 相互作用,FLO6 是水稻花粉中淀粉颗粒形成的关键成分。Western blot 结果表明,高温下 oshsp60-3b 花药中的 FLO6 水平显著降低,表明当温度超过最佳条件时,OsHSP60-3B 是稳定 FLO6 所必需的。我们认为,在高温下,OsHSP60-3B 与 FLO6 相互作用,调节水稻花粉中的淀粉颗粒生物合成,并降低花药中的 ROS 水平,以确保水稻雄配子体正常发育。

自主机器人的收敛性有限
Abstract A distributed algorithm A solves the Point Convergence task if an arbitrarily large collection of entities, starting in an arbitrary configuration, move under the control of A to eventually form and thereafter maintain configurations in which the separationbetweenallentitiesisarbitrarilysmall.Thisfundamentaltaskinthestandard O BLOT modelofautonomousmobile entities has been previously studied in a variety of设置,包括完整的可见性,确切的测量(包括距离和角度)以及实体的同步激活。我们的研究涉及最小的假设,在这些假设下,可以保证以这种方式融合的实体,具有有限和未知的可见度范围,可见度范围有限且不明显不精确。我们提出了一种在这些约束下运行的算法,该算法解决点收敛,对于在两个或三维空间中移动的实体,并具有任何有限程度的异步。我们还证明,在类似的逼真的约束下,但无限的异步,通常不可能在平面中的点收敛,这是基于自然假设,即算法在初始配置中维持存在的实体之间维持(可见的)连接性。我们称这种变体称为凝聚力融合,可以区分自主移动实体控制的有限和无限异步的力量,解决了一个长期存在的问题,一个长期存在的问题是否同步安排的实体是否比异步计划更强大。

DNA模拟的空间控制的蛋白水解靶向Cyclind1-CDK4/6复合蛋白降解的嵌合体
图2。距离和方向依赖性的选择性和Cyclin D1-CDK4/6复合物的降解。(a)DTAC距离库的示意图。(b)Western印迹(WB)分析显示,在用指定的DTAC变体以指定浓度处理的U-251细胞中细胞周期蛋白D1和CDK4/6降解14小时。结果代表了三个独立实验。(c)与距离依赖性DTAC治疗后对照组相比,相对细胞周期蛋白D1,CDK4和CDK6水平的定量。显示的数据是三个独立实验的平均值±SD。(d)DTAC方向库的示意图。(e)WB分析显示,在指定浓度的U-251细胞中,用指定的方向变体(DTAC-V5至DTAC-V9)处理的U-251细胞中的细胞周期蛋白D1和CDK4/6降解,持续14小时。结果代表了三个独立实验。(f)与对照组相比,用方向依赖性DTAC进行对照组进行了相对细胞周期蛋白D1,CDK4和CDK6水平的定量。显示的数据是三个独立实验的平均值±SD。

赖氨酸乙酰转移酶Kat8
摘要:KAT8是一种赖氨酸乙酰转移酶,主要催化组蛋白H4(H4K16)的Lys16的乙酰化。KAT8失调与许多癌症类型的发展和转移有关,包括非小细胞肺癌(NSCLC)和急性髓样白血病(AML)。到目前为止,很少有KAT8抑制剂报道,其中没有一个显示选择性活动。基于KAT3B/KDAC抑制剂C646,我们开发了一系列N-苯基-5-吡唑酮衍生物,并将化合物19和34鉴定为低微摩尔KAT8抑制剂在KAT和KDAC面板上选择性的低微球Kat8抑制剂。Western印迹,免疫荧光和CETSA实验表明,这两种抑制剂均选择性地靶向细胞中的Kat8。19和34在包括NSCLC和AML在内的不同癌细胞系中表现出Microl摩尔抗增生活性,而不会影响非转化细胞的生存能力。总体而言,这些化合物是阐明Kat8生物学的宝贵工具,它们的简单结构使它们成为有希望的未来优化研究的候选人。
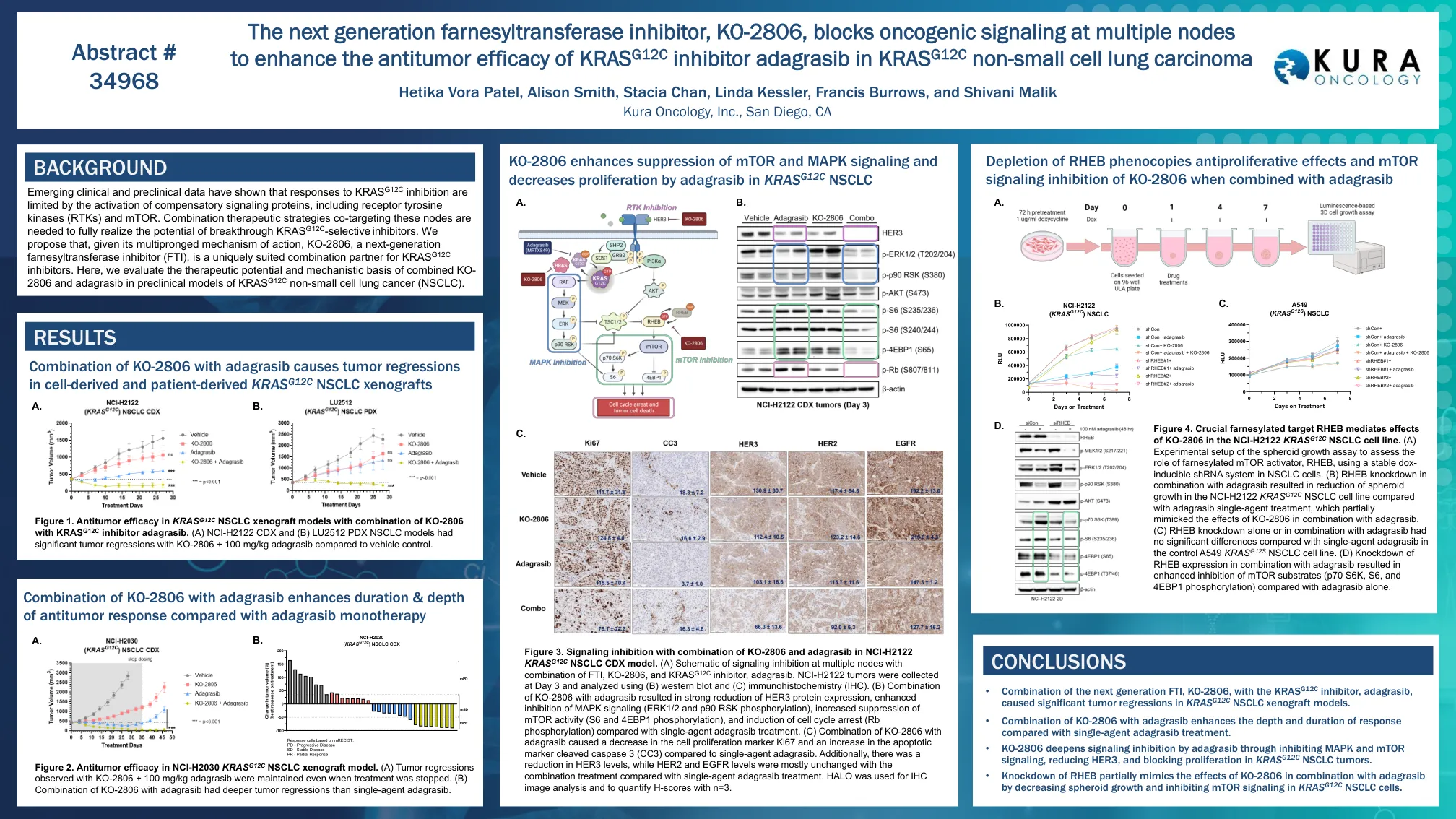
下一代Farnesylsylansferase抑制剂KO-2806,阻止多个节点的致癌信号传导,以增强KRAS G12的抗肿瘤功效
图3。NCI-H2122 KRAS G12C NSCLC CDX模型中KO-2806和Adagrasib结合使用的信号传导抑制。(a)在多个节点上抑制信号传导的示意图与FTI,KO-2806和KRAS G12C抑制剂Adagrasib的组合。NCI-H2122肿瘤,并使用(b)Western印迹和(c)免疫组织化学(IHC)分析。(b)KO-2806与AdagrasiB的组合导致HER3蛋白表达的大幅度降低,增强了对MAPK信号传导(ERK1/2和P90 RSK磷酸化)的抑制作用,增加了MTOR活性的抑制(S6和4EBP1磷酸化),以及对单个Cell Cycer Cyprant(RB Pranscratib)的抑制。(c)KO-2806与Adagrasib的组合导致细胞增殖标记Ki67的降低,而与单药adagrasib相比,凋亡标记裂解的caspase 3(CC3)的增加。此外,与单药Adagrasib治疗相比,HER3水平降低了,而HER2和EGFR水平的组合处理大多是没有变化的。halo用于IHC图像分析,并用n = 3量化H得分。

DNA 聚合酶μ兔多克隆抗体
产品名称 DNA Pol μ 兔多克隆抗体 宿主物种 兔 应用 WB;ELISA 物种交叉反应 人;大鼠;小鼠; 建议稀释度 Western Blot:1/500 - 1/2000。ELISA:1/40000。尚未在其他应用中测试。 免疫原 来自 DNA Pol μ 的合成肽。AA 范围:210-290 特异性 DNA Pol μ 多克隆抗体检测内源水平的 DNA Pol μ 蛋白。 制剂 含有 50% 甘油、0.5% BSA 和 0.02% 叠氮化钠的 PBS 液体。 储藏 储存于 -20°C。避免反复冻融循环。 蛋白质名称 DNA 指导的 DNA/RNA 聚合酶 mu 基因名称 POLM 细胞定位 细胞核。 纯化 使用表位特异性免疫原,通过亲和层析法从兔抗血清中亲和纯化抗体。克隆性 多克隆 浓度 1 mg/ml 观察到的条带 54kD 人类基因 ID 27434 人类 Swiss-Prot 编号 Q9NP87 别名 POLM;polmu;DNA 引导的 DNA/RNA 聚合酶 mu;Pol Mu;末端转移酶 背景催化活性:脱氧核苷三磷酸 + DNA(n) = 二磷酸 + DNA(n+1)。,辅因子:镁。,功能:似乎充当 Ig 变位酶,负责免疫球蛋白 (Ig) 基因超突变。,相似性:属于 DNA 聚合酶 X 型家族。,相似性:包含 1 个 BRCT