XiaoMi-AI文件搜索系统
World File Search Systemelegans

2013年秀丽隐杆线虫会议组织委员会
开幕式联谊会,6 月 26 日,星期三:10:00 至 11:30 的全体会议 1 结束后,立即在 Royce Quad 的开幕式联谊会上与朋友和同事见面。将提供免费啤酒、葡萄酒和苏打水。GSA 教师指导社交活动将于 6 月 27 日星期四晚上 7:00 至 8:00 在 Grand Horizon Ballroom Terrace 举行。本次活动由资深 C. elegans 教师为现任和新聘用的初级教师组织。将介绍新教师,然后进行非正式讨论。鼓励所有 C. elegans 实验室负责人参加。晚间社交活动,6 月 27 日星期四和 6 月 28 日星期五:晚间社交活动将于晚上 10:00 至午夜 12:00 在 Pauley Pavilion 大厅举行。将提供免费啤酒、葡萄酒和苏打水。饮料必须留在大厅。海报展示区不允许饮用任何饮料。周六晚间活动,6 月 29 日:计划于晚上 6:30 至晚上 8:00 在 Royce Quad 举行烧烤活动。C. elegans 艺术展颁奖典礼将于晚上 8:15 在 Royce Hall 举行,随后是 Worm Variety Show,时间为晚上 8:30 至晚上 9:00。会议派对将于晚上 9:00 至晚上 11:30 在 Ackerman Grand Ballroom 举行。届时将提供啤酒、葡萄酒和软饮料。用餐时间

秀丽隐杆线虫十年来的 CRISPR-Cas Gnome 编辑
摘要:CRISPR-Cas 使我们能够以前所未有的效率引入所需的基因组编辑,包括突变、表位和缺失。在模式生物的帮助下,CRISPR-Cas 的发展已经取得了很大进展,现已应用于各个领域。秀丽隐杆线虫是过去十年中迅速建立起众多 CRISPR-Cas 策略的先锋动物之一。具有讽刺意味的是,众多方法的出现使得选择正确的方法变得困难。选择合适的选择或筛选方法是规划基因组修饰的第一步。本报告总结了使用秀丽隐杆线虫的 CRISPR-Cas 方法的主要特征和应用,说明了关键策略。我们对 CRISPR-Cas 重大进展的概述将帮助读者了解基因组编辑的最新进展并浏览各种 CRISPR-Cas 基因组编辑方法。

对秀丽隐杆线虫蛋白质蛋白质的全面调查...
抽象的Argonaute(AGO)蛋白与小RNA相关,以指导其效应子在互补的转录本上。线虫秀丽隐杆线虫含有扩展的19个功能AGO蛋白质,其中许多尚未充分表征。在这项工作中,我们使用CRISPR-CAS9基因组编辑进行系统地分析了每一个秀丽隐杆线虫,以介绍GFP :: 3xflag标签。我们已经表征了整个发育过程中AGO的表达模式,鉴定了小的RNA结合补充,并确定了AGO丢失对小RNA种群和发育表型的影响。我们的分析表明将AGO的亚集分层分为不同的调节模块,并且数据的整合使我们发现了由于AGO损失而导致的新型应激诱导的生育能力和病原体反应表型。

频道介导秀丽隐杆线虫中的鼻子触摸反应
在触摸受体,胶质细胞和辅助细胞中起关键作用。然而,这种调节的基础机制知之甚少。我们首次表明,在秀丽隐杆线虫鼻触摸受体的神经胶质中需要氯化物通道CLH-1,以进行触摸反应和调节兴奋性。使用体内Ca 2+和Cl-成像,行为测定以及遗传和药理操作的组合,我们表明CLH-1介导了胶质GABA抑制灰分感官神经元功能以及用于调节灰神经元cAMP水平的CL-通量。最后,我们表明大鼠CLC-2通道挽救了CLH-1的鼻子触摸不敏感的表型,强调了整个物种功能的保护。我们的工作将神经胶质Cl-通道视为触摸灵敏度的新型调节剂。我们提出,Glial CLH-1调节Ca 2+与Ash神经元中CAMP信号之间的相互作用,以控制蠕虫的鼻子触摸受体的灵敏度。
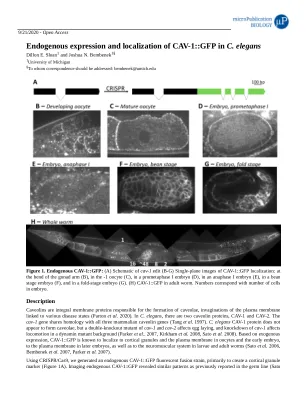
Cav-1 :: GFP在C. exkemans中的内源表达和定位
小窝蛋白是负责形成口洞的整体膜蛋白,与各种疾病状态相关的质膜的内陷(Parton等人2020)。在秀丽隐杆线虫中,有两个小窝蛋白Cav-1和cav-2。CAV-1基因与所有三个哺乳动物小窝蛋白基因共享同源性(Tang等人1997)。 秀丽隐杆线虫Cav-1蛋白似乎并不形成小窝,但是Cav-1和Cav-2的双敲击突变体会影响产卵,而Cav-1的敲低会影响动态突变体背景中的运动(Parker等人 。 2007,Kirkham等。 2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。1997)。秀丽隐杆线虫Cav-1蛋白似乎并不形成小窝,但是Cav-1和Cav-2的双敲击突变体会影响产卵,而Cav-1的敲低会影响动态突变体背景中的运动(Parker等人。2007,Kirkham等。 2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2007,Kirkham等。2008,Sato等。 2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2008,Sato等。2008)。 基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。 2006,Bembenek等。 2007,Parker等。 2007)。2008)。基于外源表达,Cav-1 :: GFP众所周知,众所周知,卵形颗粒和质膜中的质膜和早期胚胎,在后来的胚胎中,质膜,以及幼虫和成人蠕虫中的神经肌肉系统(Sato等)(Sato等。2006,Bembenek等。 2007,Parker等。 2007)。2006,Bembenek等。2007,Parker等。 2007)。2007,Parker等。2007)。2007)。

祖先糖蛋白激素受体途径控制秀丽隐杆线虫的生长
在脊椎动物中,甲状腺纤维蛋白是一种高度保守的糖蛋白激素,除了甲状腺刺激激素(TSH)外,它是TSH受体的有效配体。甲状腺激素被认为是其亚基GPA2和GPB5的最祖先糖蛋白激素和直系同源物,在脊椎动物和无脊椎动物中广泛保守。与TSH不同,甲状腺纤维蛋白神经内分泌系统的功能在很大程度上尚未探索。在这里,我们在秀丽隐杆线虫中确定了功能性甲状腺抑制蛋白样信号传导系统。我们表明,GPA2和GPB5的直系同源物以及甲状腺激素释放激素(TRH)相关的神经肽构成了促进秀丽隐杆线虫生长的神经内分泌途径。GPA2/GPB5信号是正常体型所必需的,并通过激活糖蛋白激素受体直立型FSHR-1来起作用。秀丽隐杆线虫GPA2和GPB5在体外增加了FSHR-1的cAMP信号传导。两个亚基均在肠神经元中表达,并通过向其神经胶质细胞和肠受体发出信号来促进生长。受损的GPA2/GPB5信号传导导致肠腔腹胀。此外,缺乏甲基抑制蛋白的信号传导的突变体显示出增加的排便周期。我们的研究表明,甲状腺激素GPA2/GPB5途径是一种古老的肠神经内分泌系统,可调节Ecdysozoans的肠道功能,并且可能在祖先中参与了对生物生长的控制。

秀丽隐杆线虫基因调控等位基因和记者抨击研究
基因调控是多细胞生物的重要过程,但识别功能性调控序列和机制可能具有挑战性。在秀丽隐杆线虫中,正向遗传学可以识别破坏生理过程的内源性突变(“等位基因”),从而以无偏见的方式定义功能序列(Brenner 1974;Trent、Wood 和 Horvitz 1988;Desai 等人 1988;Barton、Schedl 和 Kimble 1987)。基于 CRISPR 的基因组编辑可用于测试内源序列的功能和生理作用(Dickinson 和 Goldstein 2016;Vicencio 和 Cerón 2021)。报告基因检测中对非编码 DNA 进行系统性测试(例如“报告基因抨击”)可以识别功能序列,但不能直接检查生理功能(Aamodt、Chung 和 McGhee 1991;Didiano 和 Hobert 2006;Boulin、Etchberger 和 Hobert 2006;Nance 和 Frøkjær-Jensen 2019)。

线虫秀丽隐杆线虫:研究免疫学和微生物学的方便模型
Caenorhabditis秀丽隐杆线虫是一种线虫,在世界各地的各种环境中自然存活。该线虫已被用作发展,癌症和衰老的模型系统,因为它与人类的关键基因和疾病中涉及的信号通路共享。此外,该线虫在实验室中易于维持,并且在遗传上是可探讨的。与秀丽隐杆线虫有关的与人类癌,先天免疫和寿命相关的主要发现,但该线虫尚未用于研究与口腔健康相关的基因或微生物群。世界上几个实验室开始研究肠道菌群对秀丽隐杆线虫健康的影响。我们建议通过喂养从人类唾液中分离出的线虫细菌来研究口腔菌群对秀丽隐杆线虫的影响。能够通过秀丽隐杆线虫分析的数据来表征人口腔菌群,可以提供一种方便的方法来筛查不同口腔细菌的快速影响,并可以为几种口腔疾病提供新的前景。

nhr-49 突变影响秀丽隐杆线虫对耶尔森氏菌生物膜的敏感性
秀丽隐杆线虫转录因子 NHR-49 因其在调节代谢过程、应激反应、先天免疫和衰老方面的作用而受到广泛研究。一种以前称为 bah-3 的基因的分子鉴定表明,bah-3 ( dc9 ) 是 nhr-49 的赭色无义等位基因,该基因影响蠕虫对耶尔森氏菌细菌生物膜有害表面附着的敏感性。nhr-49 的其他严重突变也有 Bah 表型,但影响该基因 5' 同工型的缺失并不影响生物膜附着,3' 获得功能错义突变也不影响。其他 bah 基因(bah-1、bah-2、bah-4)编码 GT92 糖基化因子,预计会影响表面涂层。 NHR-49 可能充当一个或多个表面糖基化基因的正转录因子,与其在调节代谢过程中的其他作用相反。

研究秀丽隐杆线虫线粒体形态的新型成像工具
线粒体的结构和功能之间存在密切的相互作用。要理解这种复杂的关系,需要先进的成像技术来捕捉线粒体的动态特性及其对细胞过程的影响。然而,大部分关于线粒体动力学的研究都是在单细胞生物或体外细胞培养中进行的。在这里,我们介绍了一种用于实时成像秀丽隐杆线虫线粒体形态的新型遗传工具,以满足研究活体完整多细胞生物内细胞器动力学的先进技术的迫切需求。通过全面的分析,我们将我们的工具与现有方法直接进行比较,展示它们在可视化线粒体形态方面的优势,并对比它们对生物体生理学的影响。我们揭示了传统技术的局限性,同时展示了我们的方法的实用性和多功能性,包括内源性 CRISPR 标签和异位标记。通过提供根据实验目标选择最合适工具的指南,我们的工作推动了秀丽隐杆线虫的线粒体研究,并增强了不同成像模式的战略整合,以全面了解生物体内的细胞器动力学。