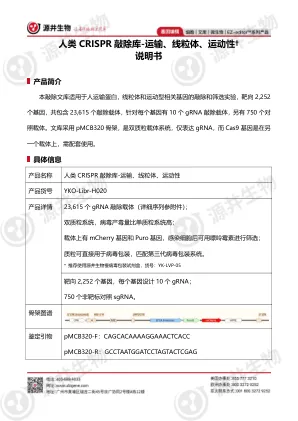
XiaoMi-AI文件搜索系统
World File Search SystemgRNA

结合工程化的 FnCas9 和扩展的 gRNA,实现 PAM 灵活、稳健且核碱基特异性的编辑和诊断
CRISPR 疗法的临床成功取决于 Cas 蛋白的安全性和有效性。来自新凶手弗朗西斯菌 (FnCas9) 的 Cas9 对错配底物的亲和力可以忽略不计,这使得它即使在结合水平上也能以非常高的精度区分 DNA 中的脱靶。然而,它的细胞靶向效率很低,限制了它在治疗应用中的使用。在这里,我们合理地设计了蛋白质以开发增强的 FnCas9 (enFnCas9) 变体,并将其细胞编辑活性扩展到以前无法访问的基因组位点。值得注意的是,一些变体释放了从 NGG 到 NGR/NRG 的原间隔区相邻基序 (PAM) 约束,使其在人类基因组位点上的可访问性增加了约 3.5 倍。enFnCas9 蛋白在体外和细胞中都具有单一错配特异性,从而扩大了基于 FnCas9 的 CRISPR 诊断的靶标范围,用于检测点突变和致病 DNA 特征。重要的是,它们在编辑效率、敲入率和脱靶特异性方面比其他经过设计的 SpCas9 高保真版本(SpCas9-HF1 和 eSpCas9)更胜一筹。值得注意的是,enFnCas9 变体可以与延长长度的 gRNA 结合使用,在 PAM 约束的规范碱基编辑器无法访问的位点进行强大的碱基编辑。最后,我们展示了使用 enFnCas9 腺嘌呤碱基编辑器完全纠正患者衍生的 iPSC 中的疾病特异性视网膜色素变性突变,突出了其在治疗和诊断中的广泛应用。

CBI-Deleted Copy Bernadette Juarez APIS 副......
然后使用“PEG 方法”将经过验证的 RNP 复合物(由单个 gRNA 和 [ ] 组成)转染到番茄原生质体中,该方法使用聚乙二醇促进 RNP 进入原生质体(Maas & Werr,1989)。进入细胞后,RNP 复合物被运输到细胞核并到达 gRNA 指示的特定目标。一旦达到目标序列,CRISPR-[ ] 酶将在 DNA 中产生双链断裂 (DSB)。当植物细胞修复断裂时,DNA 链中产生的单个断裂将重新连接,有时会导致 DNA 序列的缺失。几天后,RNP 复合物中的 CRISPR-[ ] 蛋白和 gRNA 将被植物细胞分解。

癌症中与心血管健康相关的生活质量
SLC16A2-G401R和基因敲除(KO)细胞系是通过转染重组Cas9蛋白和(G401 MUT)的合成GRNA或没有SSODN的HIPSC系列而产生的,从健康的供体(Bibhi001-b)中产生的HIPSC系。使用单个GRNA靶向外显子3生成了两种修饰(图1 A,表2)。SLC16A2-G401R Mu tation是通过使用SSODN模板通过同源性修复(HDR)引入的(图1 A,表2)。SSODN模板包括在AHDS患者中发现的突变:C.1201G> A,静音突变,以破坏PAM序列:C.1200C> T和4个无声突变,以破坏种子序列(C.1185C> T,C.1186C> A,C.1188C> A,C.1188C> A,C.1191C> a,C.1191c>1 a和f)。使用了相同的GRNA指南,并选择了带有框架移动的克隆,从而选择了过早的停止密码子。在两个报道的KO细胞系中(BIHI001-B-7和 - 8)

CRISPR 基因组编辑的最佳解决方案 - Bioneer Corporation
CRISPR(成簇的规律间隔的短回文重复序列)- Cas9 基因组编辑工具源自微生物组的适应性免疫系统,可切割目标 DNA 区域并允许 DNA 自然修复。它也被称为“RGENs”(RNA 引导的工程化核酸酶),因为它由 gRNA(向导 RNA)和 Cas9 核酸酶组成。该技术为精确操作基因组提供了更简单、更有效的方法。gRNA 由 crRNA 和 tracrRNA 组成。crRNA 具有 20 nt 的靶标互补序列,而 tracrRNA 具有 Cas9 结合所需的识别序列(图 1)。

在Cybathlon锦标赛锦标赛上竞争残疾运动员:长期运动图像脑部计算机界面训练四边形CYB
我们测试了MAD7表达质粒的共转染和GRNA表达质粒作为RNP方案的替代方案。质粒转染具有不需要先前产生和纯化MAD7蛋白的优势。我们使用与RNP实验相同的GRNA序列靶向相同的基因。如图2所示,基于质粒的协议也可以生成indels,但显示的Indels水平低于RNP协议。类似于RNP协议,尽管不太明显,但4XNLS版本优于1XNLS版本。

CRISPR指南RNA设计指南...
1.1通过CRISPR/CAS9通过特定地点的双链断裂(DSB)靶向DNA的基因组编辑(DSB)体现了现代生物技术的基石。随着新工具的发现和开发(例如工程核酸酶),对目标位点的挑战不断降低,随着时间的流逝,挑战[1]。 CRISPR/CAS9系统及其两个组件设置以及简单的目标程序构成可用工具箱中当前的金标准。 在该系统中,CAS9核酸酶的复合物和引导RNA(GRNA)介导了选定的目标位点的DSB诱导。 GRNA具有主要相关性,一方面介导靶DNA接收和结合,另一方面是Cas9的靶DNA裂解[2]。 GRNA的可变区域(指南)确定目标DNA结合的位点,并可以调整为感兴趣的序列。 采用该系统,诸如单一和多重编辑,表观遗传和转录调控,基因组基因局的可视化以及基础编辑的应用在大量生物中是可行的(有关详细信息,请参见评论[3-6])。 目标选择仅需要挑战[1]。CRISPR/CAS9系统及其两个组件设置以及简单的目标程序构成可用工具箱中当前的金标准。在该系统中,CAS9核酸酶的复合物和引导RNA(GRNA)介导了选定的目标位点的DSB诱导。GRNA具有主要相关性,一方面介导靶DNA接收和结合,另一方面是Cas9的靶DNA裂解[2]。GRNA的可变区域(指南)确定目标DNA结合的位点,并可以调整为感兴趣的序列。采用该系统,诸如单一和多重编辑,表观遗传和转录调控,基因组基因局的可视化以及基础编辑的应用在大量生物中是可行的(有关详细信息,请参见评论[3-6])。目标选择仅需要

通过将 CRISPR/Cas9 系统电穿孔到体外受精的受精卵中有效生成 GGTA1 缺陷猪
背景:异种抗原是种间异种移植成功的主要问题。GGTA1 编码 α 1,3-半乳糖基转移酶,该酶对半乳糖基-α 1,3-半乳糖的生物合成至关重要,而半乳糖是导致超急性排斥的主要异种抗原。因此,GGTA1 修饰猪是猪对人异种移植的有希望的供体。在本研究中,我们开发了一种通过电穿孔将 CRISPR/Cas9 系统引入体外受精猪受精卵以生成 GGTA1 修饰猪的方法。结果:我们设计了五种针对 GGTA1 中不同位点的向导 RNA (gRNA)。通过电穿孔将 Cas9 蛋白与每一种 gRNA 一起引入后,评估了受精卵发育成的囊胚中的基因编辑效率。使用基因编辑效率最高的 gRNA 生成 GGTA1 编辑猪。在用 Cas9/gRNA 复合物转移电穿孔受精卵后,两头受体母猪产下六头仔猪。深度测序分析显示,六头仔猪中有五头在 GGTA1 的目标区域携带双等位基因突变,没有脱靶事件。此外,用异凝集素 B4 染色证实了 GGTA1 双等位基因突变猪的 GGTA1 功能缺陷。

CRISPR 基础手册
Cas 酶是细菌免疫系统的一部分,可将短的病毒 DNA 序列整合到细菌基因组中。这是一个复杂的过程,尚未完全了解 [ 3 ]。人们所熟知的是,这些病毒序列在细菌基因组中以规则的间隔排列,彼此相距很短。这些序列之间的细菌 DNA 具有回文重复模式,因此得名,即成簇的规则间隔的短回文重复序列。整合的病毒 DNA 序列可以在需要时翻译成向导 RNA (gRNA)——也就是说,如果同一种病毒试图再次感染细菌,CRISPR 系统可以通过使用 gRNA 和 Cas 酶切割入侵的病毒 DNA。细菌免疫过程的最后一步是将 gRNA 与 Cas 结合并切割目标 DNA,这是实验室中用于基因组编辑的方法。随着 CRISPR 技术触手可及,我们进入了基因组工程的黄金时代。

1 用于多拷贝基因整合的着陆垫系统...
图 1. SD108 中全基因组整合位点的计算机筛选算法。(A)选择基因间位点中的 gRNA 进行 iCas9 介导的整合。扫描基因组中的“NGG”PAM 以获得向导 RNA 文库。筛选 gRNA 以尽量减少潜在的脱靶,并根据其基因组位置进行过滤。(B)结合各种因素对实验筛选的基因组位点进行优先排序。根据寡核苷酸合成和质粒克隆标准对 gRNA 及其相应的同源臂进行改进。实施设计规则以确保菌株稳定性,避免破坏调控元件并包括基因必需性信息,同时添加基因密度作为开放染色质的代理。结合转录组学数据来选择靠近转录活性基因的位点。