XiaoMi-AI文件搜索系统
World File Search Systemguanine
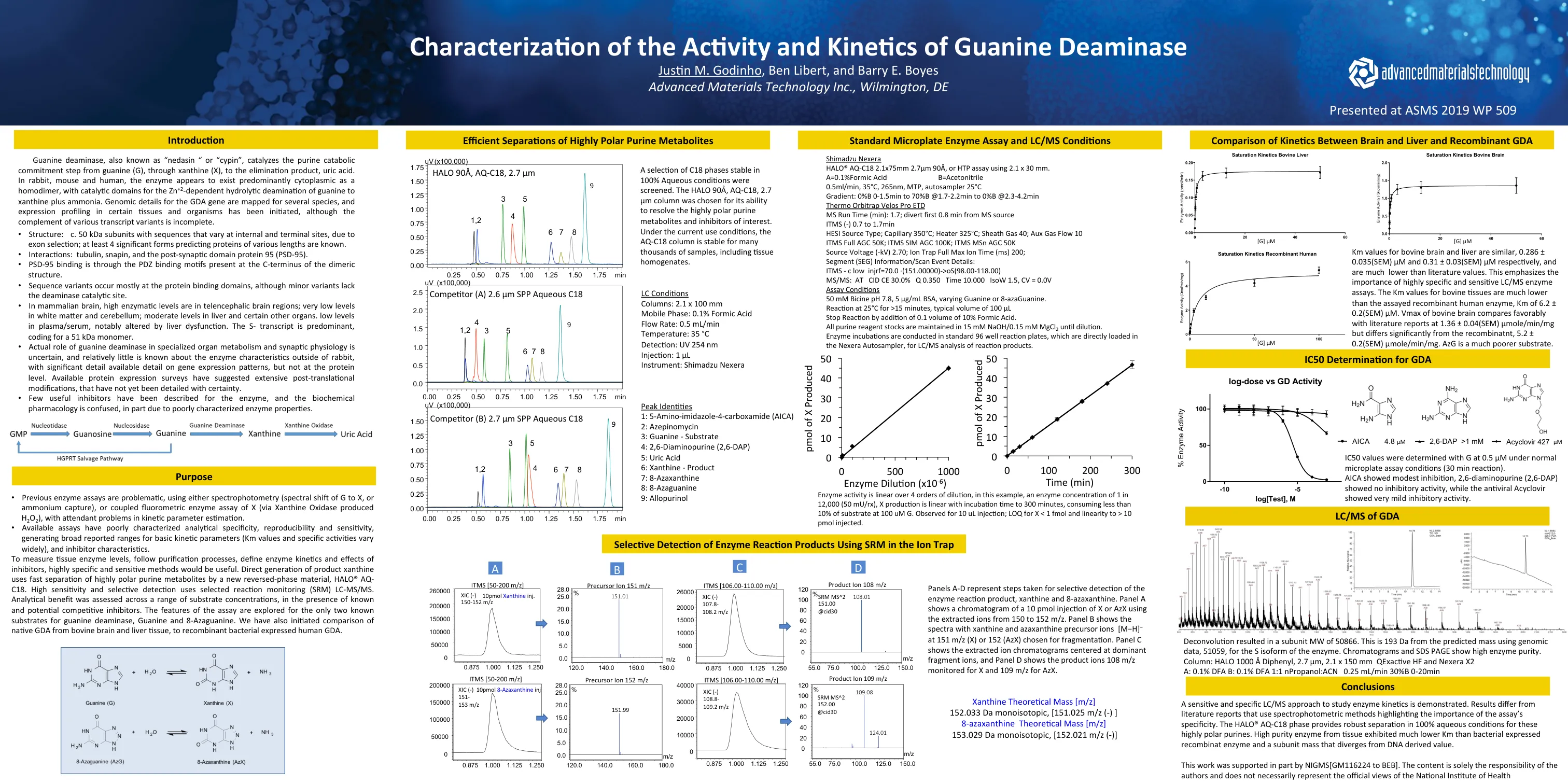
鸟嘌呤脱氨酶动力学的表征
属 要测量 直接属 高sensi 分析 对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。 我们还对来自牛脑和肝脏的Na要测量 直接属 高sensi 分析 对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。 我们还对来自牛脑和肝脏的Na直接属 高sensi 分析 对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。 我们还对来自牛脑和肝脏的Na高sensi 分析 对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。 我们还对来自牛脑和肝脏的Na分析 对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。 我们还对来自牛脑和肝脏的Na对于仅有两个已知的鸟嘌呤脱氨酶,鸟嘌呤和8-亚瓜氨酸的已知底物探索了测定的特征。我们还对来自牛脑和肝脏的Na
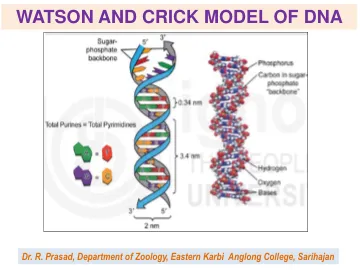
沃森和克里克的DNA模型
胸腺嘧啶和鸟嘌呤与胞嘧啶配对。腺嘌呤和胸腺嘧啶是互补碱基对。同样,胞嘧啶和鸟嘌呤也是互补碱基对。DNA的这一特性称为互补性。DNA分子中腺嘌呤的数量等于胸腺嘧啶,鸟嘌呤的数量等于胞嘧啶。腺嘌呤和胸腺嘧啶通过两个氢键连接,胞嘧啶和鸟嘌呤通过三个氢键连接。一条多核苷酸链的碱基序列决定了另一条链的碱基序列。因此,这两条链被认为是互补的。 这两条链本质上是反向平行的。一条链有3个碳

从量子透视模型看法拉第常数和蚕有什么关系?
本文尝试从量子透视模型的角度,将法拉第常数用化学核苷酸碱基(AT、G、C和U)表示。首先,将逗号后的法拉第常数的准确值排列成双数(0,96,48,53,32,12,33,10,01,84×10 5 C∙mol −1 )。其次,将这一对十进制数转换成二进制数。第三,在完成这些数的转换过程之后,再将二进制数转换成十进制数。第四,对这些十进制数分别求和。第五,将上述加法过程的总和对应到遗传密码[腺嘌呤(A)、胸腺嘧啶(T)、鸟嘌呤(G)、胞嘧啶(C)、尿嘧啶(U)]。第六,此转换的结果大致对应于尿嘧啶(U)和鸟嘌呤(G)核苷酸碱基,即数字“64”相当于尿嘧啶(U)核苷酸碱基,而近似数字“79”相当于鸟嘌呤(G)核苷酸碱基。第七,将[尿嘧啶(U)和鸟嘌呤(G)]核苷酸碱基转换为[“AG”腺嘌呤(A)和鸟嘌呤(G)]后,此结果不仅与电化学中法拉第常数之间的联系有意义,而且与量子物理学中叠加态对偶位置之间的联系也有意义。第八,在NCBI(美国国家生物技术信息中心)数据库中搜索[“AG”腺嘌呤(A)和鸟嘌呤(G)]序列后,NCBI的搜索结果与家蚕(Bombyx Mori)基因序列“AGAAAAAGGA”相似。它们是具有该序列的蚕遗传学和丝茧膜 (SCM) 基因工程可能性的非常有趣的特定模型生物。第九,这种复杂的天然蛋白质纤维膜由于具有良好的电导性而受到研究界的极大关注。最后,本文不仅揭示了法拉第常数之间的关系

电动汽车应用锂离子电池的耦合电热建模
图1。DNA结构的低能光电离已经研究了3。(a)由腺嘌呤 - 胸腺嘧啶和/或鸟嘌呤胞嘧啶碱基对组成的双链体。(b)G-四链体,其特征在于鸟嘌呤四龙的垂直堆叠(黄色);它们是由单个DNA链(单分子)的折叠,两个单链(双分子)的缔合或在含有Na +或K +阳离子(蓝色领域)的水溶液中四个单链(四分子)的关联而形成的。磷酸脱氧核糖主链以紫罗兰色指示。为简单性,在(b)中省略了环的核苷酸酶,连接鸟嘌呤四核和结束组。关于自由基阳离子的去质子化,在第3.5节中讨论了红色,蓝色和绿色质子。


DNA测序 div>
1。嘌呤碱基组(嘌呤)为太极时(胸腺素; T)和胞嘧啶; c)2。pyimidine碱基组包括腺嘌呤; A)和鸟嘌呤(G)。 Dee Syboss和磷酸盐中包括这种硝基 - 中心贝司化合物。这是由低音鲍兰与基础pyimidine(A-T,G-C)结合的一对线(图1)。所有4个核苷酸电缆的扩展都可用于分离生物可以不同的生物,使每个生物体中的遗传多样性和特异性。 div>

核酸
为了使DNA形成双链结构,重复或与RNA相互作用,碱基必须能够在一致的paxern中连接,以维持DNA序列。这涉及所谓的互补碱配对。嘌呤必须始终与嘧啶搭配,以维持相互间隔的两个分子之间的操作/mal距离。The complementary base pairing that occurs is: • The purine adenine (A) always pairs with the pyrimidine thymine (T) by forming two hydrogen bonds • The purine guanine (G) always pairs with the pyrimidine cytosine (C) by forming three hydrogen bonds • In an RNA sequence, the base thymine is replaced by uracil (U), and so this pairs with adenine 反而。


利用机器学习发现新型 SOS1 抑制剂
多种人类癌症的发病机制。1值得注意的是,KRAS 是一种常见突变,导致许多癌症病例中该基因的激活,包括 80% 至 90% 的胰腺癌、40% 至 50% 的结直肠癌和 30% 的非小细胞肺癌。1然而,对于携带 KRAS 突变的个体,临床治疗选择受到相当大的限制。目前,FDA 仅批准两种小分子抑制剂 sotorasib 和 adagrasib 用于治疗 KRAS G12C 突变的非小细胞肺癌,这表明 KRAS 靶向治疗的临床需求大大未得到满足。2,3 如图 1 所示,KRAS 的突变与 MAPK 家族中多种下游信号通路的激活有关,特别是 RAF – MEK – ERK 通路,它们对调节细胞存活和增殖至关重要。 1,4 RAS 蛋白起着分子开关的作用,在与鸟苷三磷酸 (GTP) 结合时处于活性开启状态,与与鸟苷二磷酸 (GDP) 结合时处于非活性关闭状态。5 这种开关受鸟苷酸交换因子和 GTPase 活化蛋白的调节,鸟苷酸交换因子促进 GDP 与 GTP 的交换,GTPase 活化蛋白增强 GTP 水解为 GDP。2 作为主要的鸟苷酸交换因子,Son of sevenless 1 (SOS1) 在 RAS 信号通路中起着至关重要的作用,它促进鸟苷酸交换并调节 KRAS 从“GDP 结合关闭状态”切换到“GTP 结合关闭状态”。