XiaoMi-AI文件搜索系统
World File Search Systemmilk

是什么使母乳如此惊人?
参考:1 F和Al。临床。2013; 26:29-42 EC等。 j proteom res。 2012; 11:1696-1714。 3 Genen RG,编辑。 牛奶复合材料手册。 圣地亚哥:学术出版社; 1995。 919 p。 4 Hassiotou F和Al。 细胞干。 2012; 30:2164-2174。 5 JA等。 儿科。 2001; 107:e88。 6 mouchels s和al。 临床perinatol。 2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-32 EC等。j proteom res。2012; 11:1696-1714。 3 Genen RG,编辑。 牛奶复合材料手册。 圣地亚哥:学术出版社; 1995。 919 p。 4 Hassiotou F和Al。 细胞干。 2012; 30:2164-2174。 5 JA等。 儿科。 2001; 107:e88。 6 mouchels s和al。 临床perinatol。 2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-32012; 11:1696-1714。3 Genen RG,编辑。 牛奶复合材料手册。 圣地亚哥:学术出版社; 1995。 919 p。 4 Hassiotou F和Al。 细胞干。 2012; 30:2164-2174。 5 JA等。 儿科。 2001; 107:e88。 6 mouchels s和al。 临床perinatol。 2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-33 Genen RG,编辑。牛奶复合材料手册。圣地亚哥:学术出版社; 1995。919 p。 4 Hassiotou F和Al。细胞干。2012; 30:2164-2174。5 JA等。 儿科。 2001; 107:e88。 6 mouchels s和al。 临床perinatol。 2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-35 JA等。儿科。2001; 107:e88。6 mouchels s和al。 临床perinatol。 2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-36 mouchels s和al。临床perinatol。2017; 44:193-207。 7 7找到C和Al。 br j nutr。 1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-32017; 44:193-207。7 7找到C和Al。br j nutr。1999:391-399。 8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-31999:391-399。8 Beck Kl和Al。 j proteom res。 2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-38 Beck Kl和Al。j proteom res。2015; 14:2143-2157。 9作为脱烷。 神经。 1978; 4:345-32015; 14:2143-2157。9作为脱烷。神经。1978; 4:345-3


弥合牛奶鸿沟,实现印度营养安全
牛奶消费差异 - 家庭牛奶消费 高收入群体的人均牛奶消费量是低收入群体的 3-4 倍。最贫困的 30% 家庭仅消费印度 18% 的牛奶。城市地区牛奶消费量比农村地区高 30%。拉贾斯坦邦、旁遮普邦和哈里亚纳邦每天消费超过 300 克,而奥里萨邦、恰蒂斯加尔邦和西孟加拉邦每天消费不到 171 克。获取牛奶的挑战 - 负担能力是一个关键问题;70% 的印度家庭愿意花费每月食品成本的 10-30% 来满足 300 克/天的摄入量。富裕的城市家庭过度消费,导致肥胖和健康问题。表列部落等社会群体的牛奶消费量最低。牛奶中的常量营养素和微量营养素 - 酪蛋白和乳清等优质蛋白质有助于肌肉发育和修复。钙对骨骼健康和预防骨质疏松症至关重要。维生素 A、D、B12 促进免疫功能和能量代谢。必需脂肪酸促进大脑发育。天然糖、乳糖。


人类乳汁中未检测到 COVID-19 mRNA 疫苗
此预印本的版权所有者此版本于 2021 年 3 月 8 日发布。;https://doi.org/10.1101/2021.03.05.21252998 doi: medRxiv preprint

使用牛奶口服给予索马鲁肽和替泽帕肽
预印本(未经同行评审认证)是作者/资助者。保留所有权利。未经许可不得重复使用。此版本的版权所有者于 2024 年 12 月 29 日发布。;https://doi.org/10.1101/2024.12.28.630566 doi:bioRxiv 预印本

牛奶的中红外光谱如何有助于...
模型组预测可变最大最大SDR²CV相对RMSECV RMSECV RPDCV模型质量牛奶C4(g/dl)0.01 0.23 0.10 0.10 0.03 0.03 0.93 8%3.67 3牛奶C6(g/dl)0.01 0.01 0.01 0.16 0.16 0.07 0.02 0.02 0.02 0.02 0.91 9%3.32 3牛奶C8牛奶C8牛奶C8牛奶C8(G/DL)0.011111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111111 2011得益3牛奶C10(g/dl)0.02 0.32 0.11 0.04 0.91 9%3.37 3牛奶C12(g/dl)0.02 0.41 0.13 0.13 0.04 0.92 9%3.62 3牛奶C14(g/dl)0.05 1.05 1.20 1.20 1.20 1.20 0.45 0.45 0.13 0.13 0.13 0.15 0%0.0%0.0%0.6牛奶C14_1(dl)0.00 004 dl) 21% 1.78 5 Milk C16 (g/dL) 0.12 3.32 1.20 0.40 0.94 8% 4.18 3 Milk C16_1c (g/dL) 0.01 0.24 0.07 0.03 0.73 20% 1.91 5 Milk C17 (g/dL) 0.00 0.09 0.03 0.01 0.80 13% 2.24 4 Milk C18 (g/dL) 0.05 1.32 0.40 0.15 0.84 14% 2.51 4 Milk C18_1cis9 (g/dL) 0.08 2.69 0.76 0.29 0.95 8% 4.35 2 Milk C18_2c9c12 (g/dL) 0.00 0.17 0.06 0.02 0.72 19% 1.91 5 Milk C18_2c9t11 (g/dL) 0.00 0.14 0.03 0.02 0.74 37% 1.95 6 Milk C18_3c9c12c15 (g/dL) 0.00 0.09 0.02 0.01 0.68 22% 1.77 5 Milk Tot18_1cis (g/dL) 0.09 2.77 0.82 0.31 0.95 8% 4.58 2 Milk Tot18_2 (g/dL) 0.01 0.32 0.10 0.03 0.69 15% 1.79 5 Milk Total_C18_1 (g/dL) 0.10 2.98 0.94 0.33 0.96 7% 5.18 2 Tot18_1trans (g/dL) 0.01 0.57 0.13 0.06 0.79 21% 2.17 4 Milk Total_Trans (g/dL) 0.02 0.75 0.16 0.08 0.80 19% 2.26 4 Milk isoanteiso FA (g/dL) 0.02 0.28 0.09 0.03 0.75 14% 2.00 5 Milk Odd fatty acids (g/dL) 0.03 0.50 0.16 0.04 0.83 10% 2.41 4 Milk omega3 (g/dL) 0.00 0.11 0.03 0.01 0.66 22% 1.73 5 Milk omega6 (g/dL) 0.01 0.33 0.10 0.03 0.72 14% 1.89 5 Milk SAT FA(g/dl)0.31 6.97 2.70 0.75 0.99 3%10.22 1牛奶unsat(g/dl)0.14 3.86 3.86 1.25 0.39 0.97 5%5.75 2牛奶单fa(g/dl)(g/dl)0.12 0.12 3.42 3.42 3.42 1.08 0.35 0.35 0.35 0.30 0.77 77 13.77 13.02牛奶pufa(g/dl)dl) 2.10 4牛奶SCFA(g/dl)0.05 0.80 0.35 0.10 0.93 7%3.88 3牛奶LCFA(g/dl)0.19 4.79 4.79 1.59 0.52 0.52 0.95 7%4.52 2牛奶MCFA(G/DL)
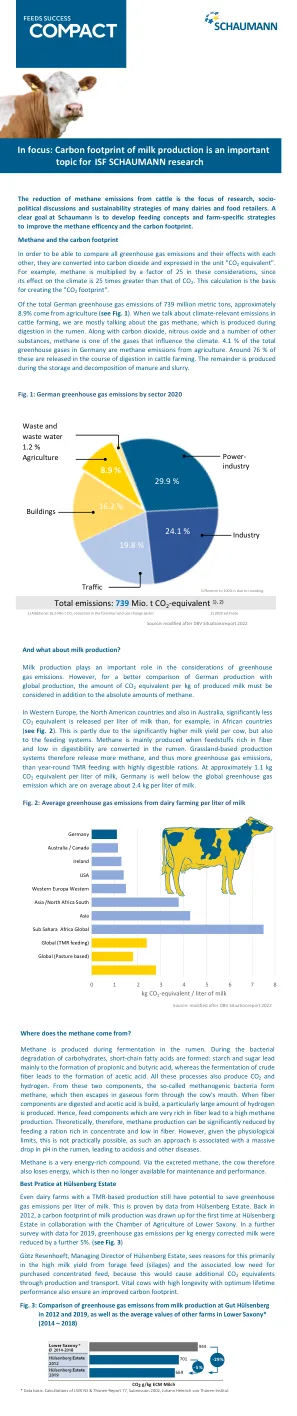
重点:牛奶生产的碳足迹很重要...
在瘤胃发酵过程中产生甲烷。在碳水化合物的细菌降解期间,形成了短链脂肪酸:淀粉和糖主要导致丙酸和丁酸的形成,而粗纤维的发酵导致乙酸的形成。所有这些过程还产生CO 2和氢。从这两个成分中,所谓的甲烷菌属形成甲烷,然后通过牛的嘴以气态逃脱。当消化纤维成分并构建乙酸时,会产生特别大量的氢。因此,富含纤维的饲料成分导致甲烷产生高。因此,从理论上讲,通过饲喂富含浓缩液和纤维低的含量可以显着降低甲烷的产生。但是,考虑到生理限制,这实际上是不可能的,因为这种方法与瘤胃的pH值大量下降有关,导致酸中毒和其他疾病。
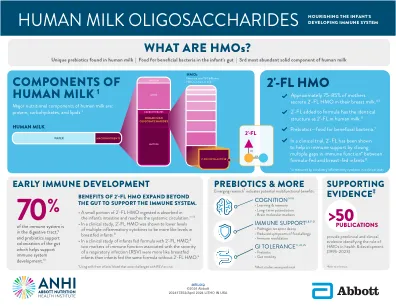
人奶寡糖滋养婴儿的
veret pa和al。J部门1995; 270:29515-29519。Erney和Al。J胃肠烯醇小儿。2000年2月; 30:181-9veret pa。食物增量J JPN。2005;(11):1018-1benenseck j和al。PLOS ONE。 2013 山和Al。 非修订版 2013年12月; 71(12):773-8 m,请参阅大胆,然后。 J农业食品化学。 2013年3月6日; 61(9):2109-1 ye,Ceng Chen,David S Newburg。 糖生物学。 2013年11月; 23(11):1281-9 chow j和al。 j proteom res。 2014; 13:2534-2542。 mcewen g和al。 Nutr Sci食物。 2014; 5:1387–1398。 kc外套和al。 PLOS ONE。 2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.PLOS ONE。2013山和Al。 非修订版 2013年12月; 71(12):773-8 m,请参阅大胆,然后。 J农业食品化学。 2013年3月6日; 61(9):2109-1 ye,Ceng Chen,David S Newburg。 糖生物学。 2013年11月; 23(11):1281-9 chow j和al。 j proteom res。 2014; 13:2534-2542。 mcewen g和al。 Nutr Sci食物。 2014; 5:1387–1398。 kc外套和al。 PLOS ONE。 2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.山和Al。非修订版2013年12月; 71(12):773-8m,请参阅大胆,然后。J农业食品化学。2013年3月6日; 61(9):2109-1ye,Ceng Chen,David S Newburg。糖生物学。2013年11月; 23(11):1281-9chow j和al。j proteom res。2014; 13:2534-2542。 mcewen g和al。 Nutr Sci食物。 2014; 5:1387–1398。 kc外套和al。 PLOS ONE。 2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2014; 13:2534-2542。mcewen g和al。Nutr Sci食物。2014; 5:1387–1398。 kc外套和al。 PLOS ONE。 2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2014; 5:1387–1398。kc外套和al。PLOS ONE。 2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.PLOS ONE。2014; 9(7):E101692。 Holscher和Al。 营养的肩膀。 2014; 144:586-5 标准A和Al。 才华。 2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2014; 9(7):E101692。Holscher和Al。营养的肩膀。2014; 144:586-5标准A和Al。才华。2014; 118:137-146。 Y,Liu s,Sun Land,DS的新闻记者。 免疫音乐。 2014; 7(6):1326-3 BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2014; 118:137-146。Y,Liu s,Sun Land,DS的新闻记者。免疫音乐。2014; 7(6):1326-3BJ的婚姻和Al。 J胃肠烯醇小儿。 2015; 61(6):649-658。 Castle-Cortin Castle L和Al。 最长。 2015; 70:1091-1 e和al。 J Nutr Biochem。 2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.BJ的婚姻和Al。J胃肠烯醇小儿。2015; 61(6):649-658。Castle-Cortin Castle L和Al。最长。2015; 70:1091-1e和al。J Nutr Biochem。2015; 26(5):455–465。 he y和al。 Nutr Adv。 2016; 7(1):102-1 kc外套和al。 j nut。 2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2015; 26(5):455–465。he y和al。Nutr Adv。2016; 7(1):102-1kc外套和al。j nut。2016; 146(12):2559-2566。 e和al。 PLOS ONE。 部分J.2016; 146(12):2559-2566。e和al。PLOS ONE。 部分J.PLOS ONE。部分J.2016; 11(11):E01666070gajzer j和al。2016; 30(补充1):671.4。 noll aj和al。 Biochem J. 2016; 473-1343-1 橄榄E,Ramire M,Vazquez E和Al。 J Nutr Biochem。 好M,Al。 br j nutr。 2016年10月; 116(7):1175–1187。 thangaram t和al。 J乳业科学。 2017; 100:7825-7833。 e和al。 br j nutr。 2017; 117(2):237-247。 高清盐和Al。 J胃肠烯醇小儿。 2017; 64(2):296-3 橄榄E和Al。 营养。 2018; 10:1 Zehra S,Cambati I和Al。 J食品科学。 2018; 83(2):499-5 ej和al。 营养。 2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;2016; 30(补充1):671.4。noll aj和al。Biochem J.2016; 473-1343-1橄榄E,Ramire M,Vazquez E和Al。J Nutr Biochem。好M,Al。br j nutr。2016年10月; 116(7):1175–1187。thangaram t和al。J乳业科学。2017; 100:7825-7833。 e和al。 br j nutr。 2017; 117(2):237-247。 高清盐和Al。 J胃肠烯醇小儿。 2017; 64(2):296-3 橄榄E和Al。 营养。 2018; 10:1 Zehra S,Cambati I和Al。 J食品科学。 2018; 83(2):499-5 ej和al。 营养。 2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;2017; 100:7825-7833。e和al。br j nutr。2017; 117(2):237-247。 高清盐和Al。 J胃肠烯醇小儿。 2017; 64(2):296-3 橄榄E和Al。 营养。 2018; 10:1 Zehra S,Cambati I和Al。 J食品科学。 2018; 83(2):499-5 ej和al。 营养。 2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;2017; 117(2):237-247。高清盐和Al。 J胃肠烯醇小儿。 2017; 64(2):296-3 橄榄E和Al。 营养。 2018; 10:1 Zehra S,Cambati I和Al。 J食品科学。 2018; 83(2):499-5 ej和al。 营养。 2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;高清盐和Al。J胃肠烯醇小儿。2017; 64(2):296-3橄榄E和Al。 营养。 2018; 10:1 Zehra S,Cambati I和Al。 J食品科学。 2018; 83(2):499-5 ej和al。 营养。 2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;橄榄E和Al。营养。2018; 10:1Zehra S,Cambati I和Al。J食品科学。2018; 83(2):499-5ej和al。营养。2018; 10:E1346。 st s和al。 2019; 63(13):E1 震惊,黄A和al。 PLOS ONE。 2019;2018; 10:E1346。st s和al。2019; 63(13):E1震惊,黄A和al。PLOS ONE。 2019;PLOS ONE。2019;

乳腺炎在牛奶质量中的重要性:文献综述
Ezequias Antonio Ortiz 1和AmanePaldêsGonçales2。 抽象的乳制品耕作是全球广泛的活动,在世界经济中占有重要空间。 目前,巴西是世界第三大牛奶生产国,每年超过340亿升。 然而,尽管具有经济和社会的重要性,但牛奶质量仍然是乳制品行业的主要挑战。 生牛奶的质量与挤奶动物的健康密切相关,这是挤奶后不久以及获得,将牛奶存储和运输到加工单元的卫生条件的初始污染程度。 在这些原因对牛奶质量造成极为有害影响的原因中,乳腺炎被认为是对乳制链影响最大的疾病。 基于该疾病在乳制品链中的重要性,本研究旨在汇编有关乳腺炎的信息,例如:其对牛奶质量,治疗和预防措施的影响。 为此,通过在线数据库中的搜索进行了调查:科学电子图书馆在线(Scielo)和PubMed,涵盖了过去五年中发表的文章。 基于关键词总共确定了175篇文章:“预防牛乳腺炎”; “牛乳腺炎治疗”; “牛乳腺炎”和“牛奶质量”。 总共包括113项研究,其中42个(37.17%)提到了预防形式,50(44.25%)用于治疗,21(18.58%)属于牛奶质量。 最后,牛奶成分和工业影响的变化加强了对乳腺炎控制的综合方法的需求。 牛乳腺炎的治疗。Ezequias Antonio Ortiz 1和AmanePaldêsGonçales2。抽象的乳制品耕作是全球广泛的活动,在世界经济中占有重要空间。目前,巴西是世界第三大牛奶生产国,每年超过340亿升。然而,尽管具有经济和社会的重要性,但牛奶质量仍然是乳制品行业的主要挑战。生牛奶的质量与挤奶动物的健康密切相关,这是挤奶后不久以及获得,将牛奶存储和运输到加工单元的卫生条件的初始污染程度。在这些原因对牛奶质量造成极为有害影响的原因中,乳腺炎被认为是对乳制链影响最大的疾病。基于该疾病在乳制品链中的重要性,本研究旨在汇编有关乳腺炎的信息,例如:其对牛奶质量,治疗和预防措施的影响。为此,通过在线数据库中的搜索进行了调查:科学电子图书馆在线(Scielo)和PubMed,涵盖了过去五年中发表的文章。基于关键词总共确定了175篇文章:“预防牛乳腺炎”; “牛乳腺炎治疗”; “牛乳腺炎”和“牛奶质量”。总共包括113项研究,其中42个(37.17%)提到了预防形式,50(44.25%)用于治疗,21(18.58%)属于牛奶质量。最后,牛奶成分和工业影响的变化加强了对乳腺炎控制的综合方法的需求。牛乳腺炎的治疗。结果表明,管理实践和新兴技术对于降低乳腺炎的发生并提高牛奶质量是基础的,而治疗替代品(例如噬菌体和纳米复合材料)则提供了有希望的溶液,以抗菌抗性抗性。关键字:预防牛乳腺炎。牛乳腺炎和牛奶质量。