XiaoMi-AI文件搜索系统
World File Search Systemtranscription
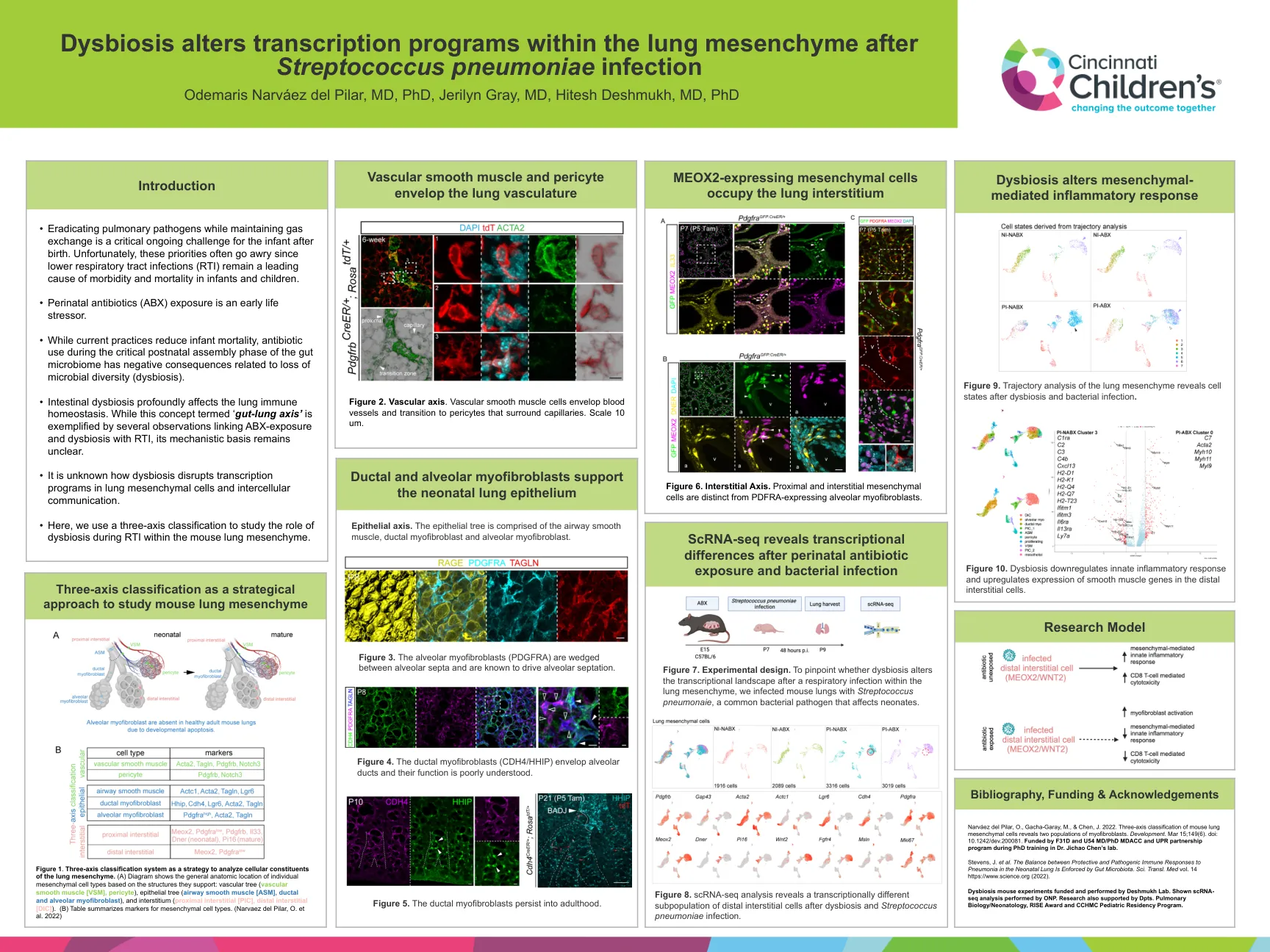
营养不良改变肺间隙内的转录程序
图1。三轴分类系统是分析肺间充质细胞成分的策略。(a)图显示了基于其支持的结构的单个间质细胞类型的一般解剖位置:血管树(血管平滑肌[VSM],周围树),上皮树(Airway平滑肌[ASM],导管和肺泡的肌肉肌纤维细胞和肌肌纤维细胞)和植物(proircimal instrastial superstitial institial softistial softsitial confiral [pic] pic](b)表总结了间充质细胞类型的标记。(Narvaez del Pilar,O。et al。2022)


人类卫星 3 DNA 编码兆碱基规模的转录因子结合平台
真核生物基因组中经常散布着大量串联重复序列,称为卫星 DNA,这些序列是组成性异染色质的基础,常位于着丝粒区域周围。虽然某些卫星 DNA 类型在着丝粒生物学中具有明确的作用,但其他丰富的卫星 DNA 的功能尚不明确。例如,人类卫星 3 (HSat3) 约占人类基因组的 2%,形成高达数十兆碱基的巨大阵列,但这些阵列在着丝粒功能中没有已知的作用,直到最近才几乎完全被排除在基因组组装之外。因此,这些巨大的基因组区域仍然相对研究不足,而 HSat3 的潜在功能作用在很大程度上仍然未知。为了解决这个问题,我们对新的 HSat3 结合因子进行了系统筛选。我们的工作表明,HSat3 阵列含有高密度的转录因子 (TF) 基序,这些基序与与多个高度保守的信号通路相关的因子结合。出乎意料的是,HSat3 中最富集的 TF 属于 Hippo 通路转录效应子家族 TEAD。我们发现 TEAD 以细胞状态特异性的方式将辅激活因子 YAP 募集到 HSat3 区域。利用 RNA 聚合酶-I 报告基因检测、HSat3 的靶向抑制、YAP 的诱导降解和超分辨率显微镜,我们表明 HSat3 阵列可以将 YAP/TEAD 定位在核仁内,YAP 在那里调节 RNA 聚合酶-I 活性。除了揭示 Hippo 通路与核糖体 DNA 调控之间的直接关系外,这项研究还表明卫星 DNA 可以编码多个转录因子结合基序,为这些巨大的基因组元素定义了新的作用。

体外转录的机械模型,结合了焦磷酸镁结晶的作用
图5代表延长的绿色荧光蛋白(EGFP),SARS -2 COV -2 SPIKE蛋白受体结合结构域(RBD)和CAS9模型验证的各种组合:IVT长度的多个DNA序列的IVT产量(Rosa等,2022)。关于模型预测的阴影区域是95%的预测间隔(SI第4节)。数据点上的误差线是根据Rosa等人的整个数据集估计的标准偏差的95%置信区间。(2022)。ivt,体外转录。Rosa等人使用的 DNA序列。 是延长的绿色荧光蛋白(EGFP),SARS -CV -2尖峰蛋白受体结合结构域(RBD)和Cas9基因的组合。DNA序列。是延长的绿色荧光蛋白(EGFP),SARS -CV -2尖峰蛋白受体结合结构域(RBD)和Cas9基因的组合。

Marchantia营养发展期间转录因子启动子活动的景观
转录因子(TFS)对于调节基因表达和细胞命运测定至关重要。表征TF基因在时空和时间上的转录活性是了解复杂生物系统的关键步骤。苔藓植物的营养植物分子分生组织具有一些特征,可以与流动植物的芽根尖分生组织具有。然而,与配子植物组织相关的TF的身份和表达方法在很大程度上尚不清楚。只有约450个假定的TF基因,马尔丁塔蒂亚(马丁坦蒂亚多形)是植物系统生物学的出色模型系统。我们已经产生了来自Marchantia TF基因的启动子元素的近乎完整的集合。我们在集合中为所有TF启动子进行了经验测试的记者融合,并系统地分析了Marchantia Gemmae中的表达模式。这使我们能够在早期营养发展中构建表达域的图,并确定一组在干细胞区域中活跃的TF衍生启动子。细胞标记提供了其他工具,并深入了解了配子分生组织的动态调节及其进化。此外,我们为集合中的所有启动子提供了在线表达模式的在线数据库。我们期望这些启动子元素将有助于细胞类型特异性,合成生物学应用和功能基因组学。

人类 3D 染色质对心脏病相关转录因子的剂量依赖性敏感性
。CC-BY-ND 4.0 国际许可证(未经同行评审认证)是作者/资助者,他已授予 bioRxiv 永久展示预印本的许可。它是此预印本的版权持有者此版本于 2025 年 1 月 12 日发布。;https://doi.org/10.1101/2025.01.09.632202 doi:bioRxiv 预印本

RFX 转录因子调节动物现存最亲属的纤毛发生
对于现代动物而言,在正确的时间在正确的细胞中部署纤毛对于发育和生理至关重要。两种转录因子 RFX 和 FoxJ1 可协调动物的纤毛发生 7–9 ,但在许多其他有纤毛的真核生物的基因组中却不存在,这引发了一个问题:动物纤毛发生的调控是如何进化的 10,11 。通过将动物的基因组与其现存最亲近的亲属领鞭毛虫的基因组进行比较,我们发现它们最后的共同祖先的基因组编码了至少三个 RFX 旁系同源物和一个 FoxJ1 同源物。模型领鞭毛虫 Salpingoeca rosetta 中 RFX 同源物 cRFXa 的破坏导致细胞增殖延迟和纤毛发生异常,以新生纤毛的崩溃和吸收为标志。在 cRFXa 突变体中,纤毛发生基因和 foxJ1 显著下调。此外,S. rosetta 纤毛基因的启动子富含与体外 cRFXa 蛋白结合的 DNA 基序相匹配的 DNA 基序。这些发现表明,祖先 cRFXa 同源物协调了动物和领鞭毛虫祖先的纤毛发生,并且选择性

旨在靶向HIF-1转录因子的抗癌剂的最新发展概述
恶性实体肿瘤迅速生长,氧气区域低于生理水平的形成。肿瘤缺氧是由于癌细胞中氧气供应与氧气消耗之间的不平衡引起的,例如与肿瘤生长相比,肿瘤脉管系统的形成速率相对较慢[1-4]引起[1-4]。癌细胞位于肿瘤血管远端区域,由于围绕血管的癌细胞的氧气消耗,导致恶性实体瘤的低氧区域,因此无法获得足够的氧气[1-4]。肿瘤脉管系统的曲折和漏水结构也是肿瘤缺氧的原因之一[1-4]。缺氧导致癌症的恶性表型和治疗性。[1 - 4]。已经揭示了细胞对缺氧的反应受某些因素的调节,但是缺氧诱导因子1(HIF-1)诱导与血管生成有关的各种基因的转录,与血管生成,葡萄糖代谢,细胞增殖,生存,入侵和转移相关,被识别为高氧响应的大师调节剂[5-7-7-7-7-7-7-7-7 ressian。HIF-1途径是预防癌症侵略性并提高癌症治疗有效性的有吸引力的目标。HIF-1是一个构成HIF-1α和HIF-1β子单位的异二聚体转录因子(图1)。HIF-1α表达受到细胞周围的氧气水平的影响,并在低氧条件下诱导。相反,构成表达的HIF-1β亚基,也称为芳基羟基受体核转运剂(ARNT)

结合IVU + OCT数据,生物力学模型和机器学习方法,用于精确冠状动脉斑块形态定量和帽厚度以及应力/应变指数预测
脊椎动物视觉系统的光感受器的发展受复杂的转录调节网络控制。otx2在有丝分裂视网膜祖细胞(RPC)中表达,并控制感光体发生。由OTX2激活的CRX在细胞周期出口后在感光前体中表达。neurod1也存在于可以指定为杆和锥形光感受器亚型中的光感受器前体中。NRL,并调节包括孤儿核受体NR2E3在内的下游杆特异性基因,该基因进一步激活了杆特异性基因并同时抑制了锥体特异性基因。锥形亚型规范也受到诸如THRB和RXRG等几个转录因子的相互作用的调节。这些关键转录因子中的突变是出生时眼部缺陷的原因,例如微感染和遗传感受器疾病,例如Leber先天性症状(LCA),色素性视网膜炎(RP)和盟友性疾病。特别是,许多突变是以常染色体主导方式遗传的,包括CRX和NRL中的大多数错义突变。在这篇综述中,我们描述了与上述转录因子中突变相关的光感受器缺陷的光谱,并总结了当前对致病突变下的分子机制的知识。终于,我们考虑了理解基因型 - 表型相关性和轮廓途径的杰出差距,以实现对治疗策略的未来研究。

植物染色质的移动:转录和 DNA 修复过程中染色质移动性的概述
近年来,人们越来越清楚地认识到染色体是高度动态的实体。染色质的移动和重排涉及许多生物过程,包括基因调控和基因组稳定性的维持。尽管对酵母和动物系统中的染色质移动性进行了广泛的研究,但直到最近,对植物中染色质移动性的研究还不多。植物要实现正常的生长和发育,就需要对环境刺激做出快速而适当的反应。因此,了解染色质移动性如何支持植物的反应可能会为植物基因组的功能提供深刻的见解。在这篇综述中,我们讨论了与植物染色质移动性相关的最新技术,包括染色质在各种细胞过程中的作用的可用技术。