XiaoMi-AI文件搜索系统
World File Search Systemtransduced
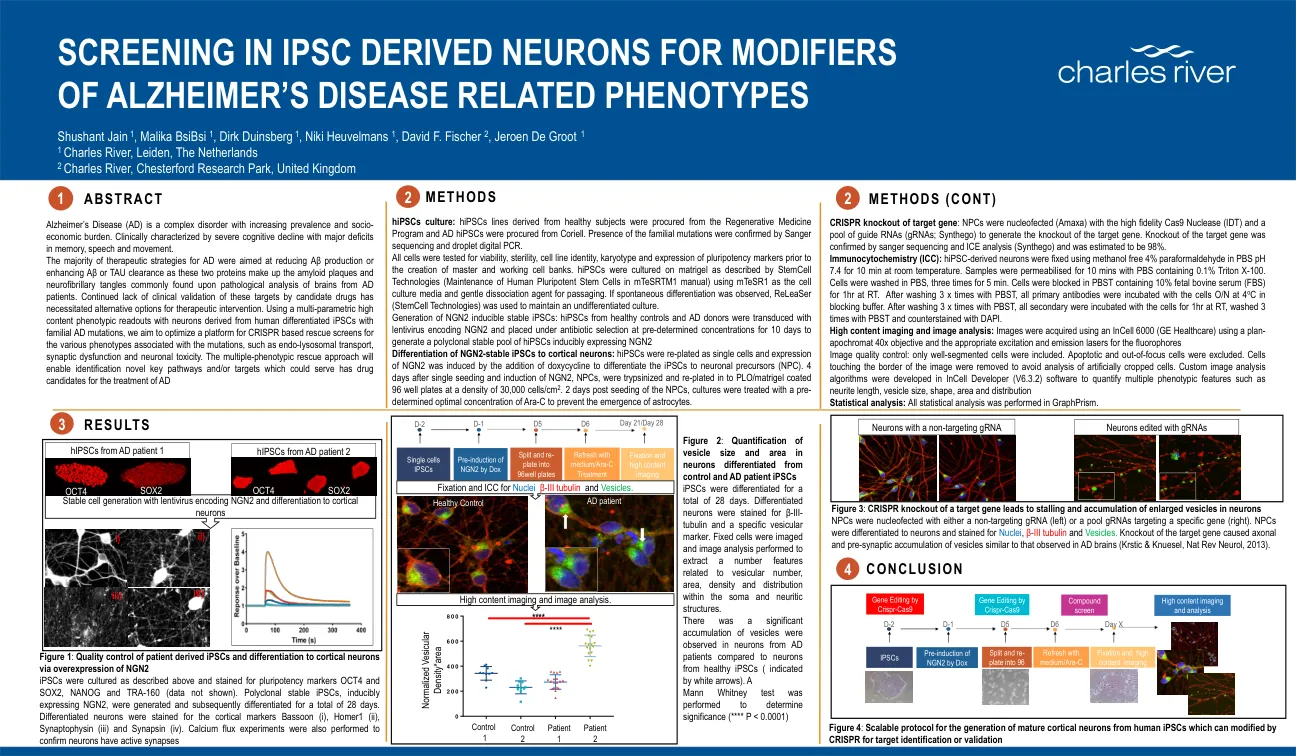
在 IPSC 衍生的神经元中筛选阿尔茨海默病相关表型的修饰因子 | Charles River
hiPSC 培养:来自健康受试者的 hiPSC 系来自再生医学计划,AD hiPSC 来自 Coriell。通过 Sanger 测序和液滴数字 PCR 确认存在家族性突变。在创建主细胞库和工作细胞库之前,对所有细胞进行了活力、无菌性、细胞系身份、核型和多能性标志物表达测试。hiPSC 在基质胶上培养,如 StemCell Technologies(mTeSRTM1 手册中的人类多能干细胞维护)所述,使用 mTeSR1 作为细胞培养基和温和的解离剂进行传代。如果观察到自发分化,则使用 ReLeaSer(StemCell Technologies)来维持未分化培养。 NGN2 诱导的稳定 iPSC 的生成:健康对照和 AD 供体的 hiPSC 被编码 NGN2 的慢病毒转导,并在预定浓度的抗生素选择下放置 10 天,以生成可诱导表达 NGN2 的 hiPSC 多克隆稳定池。NGN2 稳定的 iPSC 向皮质神经元的分化:将 hiPSC 重新接种为单细胞,并通过添加强力霉素诱导 NGN2 的表达,以将 iPSC 分化为神经元前体 (NPC)。单次接种和诱导 NGN2 4 天后,将 NPC 用胰蛋白酶消化并重新接种到 PLO/matrigel 包被的 96 孔板中,密度为 30,000 个细胞/cm 2 。在 NPC 接种 2 天后,用预定的最佳浓度的 Ara-C 处理培养物以防止星形胶质细胞的出现。

创伤性脑损伤小鼠模型中毛囊凸起衍生干细胞的多模态成像
摘要 创伤性脑损伤 (TBI) 是一种毁灭性的事件,目前治疗方法有限。干细胞移植可通过不同的机制恢复功能,例如通过分化进行细胞替换、刺激血管生成和支持微环境。成人毛囊凸起衍生干细胞 (HFBSC) 具有神经元分化能力,易于采集且相对免疫特权,这使它们成为自体干细胞治疗的潜在候选者。在本研究中,我们应用体内多模态、光学和磁共振成像技术来研究小鼠 TBI 模型中小鼠 HFBSC 的行为。HFBSC 表达 Luc2 和 copGFP,并在体外检查其分化能力。随后,在受伤 2 天后,将预装了 ferumoxytol 的转导 HFBSC 移植到裸鼠的 TBI 病变(皮质区域)旁边。移植后 58 天将大脑固定以进行免疫组织化学检测。表达 Luc2 和 copGFP、载有 ferumoxytol 的 HFBSC 在体外表现出足够的神经元分化潜能。受损大脑的生物发光显示 HFBSC 存活,磁共振成像确定了它们在移植区域的定位。免疫组织化学显示移植细胞染色为巢蛋白和神经丝蛋白 (NF-Pan)。细胞还表达层粘连蛋白和纤连蛋白,但未检测到细胞外基质团块。58 天后,可以在脑组织切片中的 HFBSC 中检测到 ferumoxytol。这些结果表明 HFBSC 能够在脑移植后存活,并表明细胞可能向神经元细胞谱系分化,这支持了它们在 TBI 细胞治疗中的潜在用途。

CRISPR/Cas9 基因敲除及随后的野生型和突变型基因拯救的改进策略
将荧光标记 mOrange 插入到流行的 pLentiCrispr-V2 中,以创建包含嘌呤霉素选择和荧光标记的 pLentiCrispr-V2-mOrange (V2mO),使病毒产生和靶标转导可见。用该质粒和适当的向导 RNA (gRNA) 包装的慢病毒成功敲除了人胃癌细胞系中的 RhoA、Gli1 和 Gal3 基因。Cas9-gRNA 编辑效率可以直接从 Cas9-gRNA 转导细胞中 gRNA 区域周围的短聚合酶链反应产物的 Sanger 电泳图来估计。必须对转导的靶细胞池进行单克隆以建立稳定的敲除克隆。仅当 gRNA 结合的 cDNA 被三个核苷酸修饰而氨基酸序列保持不变时,才能成功将野生型(RhoA 和 Gal3)和突变型(RhoA.Y42C)基因拯救到敲除细胞中。在 Gal3 基因中观察到严格的靶向 CRISPR/Cas9 编辑,但在 RhoA 基因中未观察到,因为 RhoA.Y42C 已经在 gRNA5 结合位点出现核苷酸变化。总之,我们改进的策略增加了这些优势:在流行的慢病毒系统中添加可视化标记、监测慢病毒的生产和转导效率、通过荧光激活细胞分选在靶细胞中分选 Cas9+ 细胞、通过 gRNA 结合位点周围的短 PCR 电泳图直接估计靶细胞池的基因编辑效率、以及在敲除细胞中成功拯救野生型和突变型基因,通过修饰 cDNA 克服 Cas9 编辑。

使用新的稀疏监督自动编码器神经网络
基于CRISPR的单细胞转录组筛选是有效的遗传工具,可同时评估由一组指南RNA(GRNA)靶向的细胞的表达式,并从观察到的扰动中推断靶基因函数。然而,由于各种局限性,这种方法在检测弱扰动方面缺乏灵敏度,并且在研究主调节器(例如转录因子)时基本上是可靠的。为了克服检测微妙的GRNA诱导的转录组扰动和对响应最快的细胞进行分类的挑战,我们开发了一种新的监督自动编码器神经网络方法。我们稀疏的监督自动编码器(SSAE)神经网络提供相关特征(基因)和实际扰动细胞的选择。我们将此方法应用于基于基于缺氧的长期非编码RNA(LNCRNA)的子集的基于内部单细胞CRISPR干扰(CRISPRI)转录组筛查(CROCPRI)转录组筛选(CROP-SEQ),该子集受缺氧调节的疾病,该疾病在肺腺癌(Lung adenacoarcinoma)(LUAD)的背景下促进了肿瘤的侵略性和耐药性。针对LNCRNA的子集进行了经过验证的GRNA的农作物序列库,并且作为阳性对照,HIF1A和HIF2A(低氧反应的2个主要转录因子)在3、6或24 h期间在正态氧中培养的A549 LUAD细胞中转导的2个主要转录因子。我们首先通过确定在低氧反应的时间开关期间确定其敲低的特定效应,从而验证了HIF1A和HIF2上的SSAE方法。接下来,SSAE方法能够检测出稳定的短缺氧依赖性转录组特征,该特征是由某些LNCRNA候选者的敲低诱导的,表现优于先前发表的

敲低胎盘主要的促进剂超家族域,含有2A的孕妇小鼠降低了胎儿脑的生长和磷脂docosahexaenoic Acid
摘要:简介:Docosahexaenoic Acid(DHA)是n -3长链多不饱和脂肪酸,对于胎儿发育至关重要,胎盘通过胎盘从母亲传输到胎儿。含有2A(MFSD2A)的主要促进剂超级家族型溶血磷脂酰胆碱(LPC)转运蛋白位于人胎盘的合成型胞植物细胞的基础质膜中,人胎盘的胎盘膜细胞和MFSD2A表达与人类表达的人类表达与昏迷的corn lumbilical Corncly lppc-lpc-lpc-dha相关。我们假设孕妇小鼠中MFSD2A的胎盘特异性敲低会减少胎儿脑中的磷脂DHA的积累。方法:用表达EGFP的慢病毒(E3.5)的小鼠胚泡(E3.5),该慢病毒含有靶向MFSD2A或非编码序列(SCR)的shRNA,然后转移到假孕妇中。在E18.5时,称重胎儿,并收集其胎盘,大脑,肝脏和血浆。MFSD2A mRNA表达通过QPCR在大脑,肝脏和胎盘中测定,以及通过LC-MS/MS量化磷脂DHA。结果:与SCR对照相比,在E18.5(n = 45,p <0.008)时,靶向MFSD2A的shRNA在E18.5(n = 45,p <0.008)时将胎盘mRNA MFSD2A的表达降低了38%。MFSD2a在胎儿脑和肝脏中的表达不变。胎儿脑体重减少了13%(p = 0.006)。体重,胎盘和肝脏重量不受影响。胎儿脑磷脂酰胆碱和磷脂酰乙醇胺DHA含量较低,胎盘特异性MFSD2A敲低的胎儿含量较低。这些数据提供了机械证据,表明胎盘MFSD2A介导LPC-DHA的母体 - 饮食转移,这对于大脑生长至关重要。结论:LPC-DHA转运蛋白MFSD2A表达表达的胎盘特异性减少导致胎儿脑体重降低,胎儿大脑中磷脂DHA含量降低。

开展 CHO DG44 全基因组 KO 筛选,揭示基因组学与快速生长表型之间的联系
中国仓鼠卵巢 (CHO) 细胞系广泛应用于生物制药生产。细胞系生成的改进加快了最终生产克隆的速度,但开发新型生物分子、生产力限制和市场需求方面的挑战使得细胞系开发 (CLD) 必须不断改进。虽然细胞生长在 CLD 期间显示出明显的瓶颈,但对 CHO 细胞系生长表型的研究有限。最近的一项研究成功地分离并永生化了一种源自原代肺细胞的新型中国仓鼠细胞系,该细胞系表现出更快的生长速度、稳定的生产力和高水平的生物制药蛋白质生产 1 。值得注意的是,CHL-YN 细胞系的倍增时间缩短至 10.7 小时,而 CHO 细胞系的倍增时间通常为 18.0 至 22.0 小时。在这里,我们旨在进行全面的全基因组敲除 (KO) 筛选,以确定加速 CHO 细胞生长的遗传靶点和途径,揭示与 CHO 细胞生长相关的基本遗传机制。我们建立了一个强大的 CRISPR 能力的 CHO DG44 细胞系,能够在单向导 RNA (sgRNA) 存在的情况下以可预测的方式产生插入/删除 (InDel) 事件。此外,我们测试了使用小型 140 sgRNA 微型文库生成和培养转导文库的方法。我们优化的设置能够实现约 80% 的单拷贝整合,这比最近文献中的过去工作有所改进 2 。此外,我们为影响生长的基因靶标的 CRISPR 核酸酶表达依赖性富集和消耗效率提供了证据。

lyfgenia®(lovotibeglogene autotemcel)
严重影响的患者可能会经历多种并发症,例如由于小周围障碍物(Vaso-Occlyclusive Carlisis [voc];镰状细胞危机),急性疾病,急性胸部综合征(ACS;与肺炎症状的急性症状)有关,由于小血管造成的疾病(VOS-cocle危机),反复发生的急性疼痛(VOE)反复急性疼痛(VOE)(VOES),可能是症状,刺激性症状。骨骼,肾脏,心脏,肝脏和肺部或导致严重的感染并发症,例如功能性低下和早亡。治疗包括控制并发症,缓解疼痛,防止感染并最大程度地减少器官损伤的措施。标准药理治疗包括药物,例如羟基脲(HYDEREA),镇痛药和输血。造血干细胞移植适当的供体患者,直到基因治疗的发展为止一直是治愈的选择。基因疗法现在为没有愿意HLA匹配的家庭捐助者的严重镰状细胞疾病的成员提供治疗选择。食品药物管理局(FDA)批准的适应症:•Lyfgenia是一种自体造血干细胞基因疗法,用于治疗12岁或以上患有镰状细胞疾病的患者和血管合作术的病史。lyfgenia使用过体内慢病毒载体基因疗法,该疗法通过通过BB305 LVV。HBAT87Q具有相似的氧气 -输注Lyfgenia后,转导的CD34+ HSC植入了骨髓中并分化以产生含有生物活性βA-T87Q-珠蛋白的红细胞,该细胞将与α-蛋白结合起来,从而产生含有βA-T87Q-球蛋白(HBAT87Q)的功能性HB。

T细胞受体T细胞(TCR T)治疗
收养细胞疗法(ACT)使用基因工程的人类淋巴细胞,并且正在越来越多地研究血液系统恶性肿瘤和实体瘤的患者。采取两种方法包括嵌合抗原受体(CAR)T细胞疗法,该疗法使用引入免疫效应细胞的人造受体识别肿瘤细胞表面抗原和T-cell受体(TCR)基于基于遗传修饰的淋巴细胞的肿瘤细胞表面抗原(TCR)受体(TCR),这些疗法针对特定的肿瘤标记。基于TCR的治疗涉及患者筛查,白细胞术,转导的TCR产物的产生,淋巴结序列以及基于TCR的产物的输注。基于TCR的疗法的挑战包括与淋巴结蛋白的产品制造,患者选择和制备相关的疗法。TCR的治疗越来越多地研究了实体瘤的治疗,因为它可以识别由肿瘤细胞表面上主要的组织相容性复合物(MHC)分子提出的肿瘤特异性表位。该策略具有可能更广泛的适用性,因为细胞内的肿瘤特异性序列比表面上的肿瘤特异性蛋白要多得多。这些细胞内癌靶标仅通过基于TCR的方法而不是通过基于CAR的方法来访问(Tsimberidou等,2021)。原则上,ACT可以使用各种效应细胞,但最常见的是基于T细胞或天然杀伤(NK)细胞,该细胞源自患者并进行了遗传修饰。大多数免疫疗法失败,因为它们无法提供有效的抗肿瘤效应细胞池和/或因为动员的效应细胞受到肿瘤相关因子的抑制。基于TCR的治疗通过体内生产具有已知选择性和效力的活化淋巴细胞的生产(Ping等,2018)。

BAd-CRISPR:在成年小鼠肩胛间棕色脂肪组织中诱导基因敲除
CRISPR/Cas9 已实现多种组织中的可诱导基因敲除;然而,尚未有其在棕色脂肪组织 (BAT) 中的应用报道。在此,我们开发了棕色脂肪细胞 CRISPR (BAd-CRISPR) 方法来快速检测一个或多个基因的功能。使用 BAd-CRISPR,将表达单向导 RNA (sgRNA) 的腺相关病毒 (AAV8) 直接施用于在棕色脂肪细胞中表达 Cas9 的小鼠的 BAT。我们表明,将 AAV8-sgRNA 局部施用于成年小鼠的肩胛间 BAT 可强有力地转导棕色脂肪细胞,并使脂联素、脂肪甘油三酯脂肪酶、脂肪酸合酶、周脂素 1 或硬脂酰辅酶 A 去饱和酶 1 的表达降低 90% 以上。施用多个 AAV8 sgRNA 可同时敲除多达三个基因。 BAd-CRISPR 诱导移码突变并抑制靶基因 mRNA 表达,但不会导致 BAT 中脱靶突变的大量积累。我们利用 BAd-CRISPR 创建了可诱导的解偶联蛋白 1 (Ucp1) 敲除小鼠,以评估 UCP1 缺失对成年小鼠适应性产热的影响。可诱导的 Ucp1 敲除不会改变核心体温;然而,BAd-CRISPR Ucp1 小鼠的成纤维细胞生长因子 21 循环浓度升高,并且 BAT 基因表达发生变化,与通过增加过氧化物酶体脂质氧化而产生的热量一致。其他分子适应性预示着额外的细胞效率低下,蛋白质合成和周转增加,线粒体对线粒体编码基因表达的依赖降低,核编码线粒体基因表达增加。这些数据表明 BAd-CRISPR 是一种加速脂肪组织生物学发现的有效工具。

choe_watchmaker_simic_2021.pdf - Lim 实验室 | UCSF
使用嵌合抗原受体 (CAR) T 细胞治疗实体癌的难题是缺乏理想的靶抗原,这些靶抗原既绝对肿瘤特异性又均质表达。我们表明,多抗原引发和杀伤识别电路提供了灵活性和精确度,可以克服胶质母细胞瘤背景下的这些挑战。可识别特定引发抗原(例如异质但肿瘤特异性的胶质母细胞瘤新抗原表皮生长因子受体剪接变体 III (EGFRvIII) 或中枢神经系统 (CNS) 组织特异性抗原髓鞘少突胶质细胞糖蛋白 (MOG))的 synNotch 受体可用于局部诱导 CAR 表达。通过靶向同质但非绝对肿瘤特异性的抗原,这可以彻底但可控地杀死肿瘤细胞。此外,synNotch 调节的 CAR 表达可避免强直信号和衰竭,使更高比例的 T 细胞保持幼稚/干细胞记忆状态。在携带脑内患者来源异种移植瘤 (PDX) 且异质性表达 EGFRvIII 的免疫缺陷小鼠中,单次静脉输注 EGFRvIII synNotch-CAR T 细胞表现出比传统组成性表达 CAR T 细胞更高的抗肿瘤功效和 T 细胞持久性,且不会杀死肿瘤外细胞。用中枢神经系统特异性抗原 MOG 引发的 synNotch-CAR 电路转导的 T 细胞也表现出对脑内 PDX 的精确和有效控制,而没有证据表明在脑外引发。总之,通过使用整合对多个不完善但互补抗原的识别的电路,我们提高了针对胶质母细胞瘤的 T 细胞的特异性、完整性和持久性,从而提供了适用于其他实体肿瘤的通用识别策略。